Lexikon der Biologie: Symmetrie
ESSAY
Peter Sitte
Symmetrie
Der Symmetriebegriff
Symmetrie [von griech. symmetria = Ebenmaß] ist die geordnete Wiederholung gleicher Strukturelemente ( vgl. Infobox ). Sie wird überall dort beobachtet, wo es einander zugeordnete, aufeinander bezogene Ähnlichkeiten gibt. Symmetrie äußert sich im Auftreten regelmäßiger Muster. Die Musterelemente können gedanklich durch sog. Deckoperationen aufeinander abgebildet werden. So wird auf einer mit lauter E gleichmäßig bedruckten Seite ein willkürlich herausgegriffenes E bei Verschiebung in Zeilenrichtung oder senkrecht dazu immer wieder mit anderen E zusammentreffen und diese „decken“.
Die meisten Organismen lassen auffällige Symmetrien erkennen. Eine mathematische Behandlung von Biosymmetrien ist allerdings weniger sinnvoll als bei Kristallgittern (Kristall), und die Symmetriekriterien müssen in der Biologie weiter gefaßt werden als in der Kristallographie (Proteinkristallisation, Röntgenstrukturanalyse). Wesentlich ist aber auch in der biologischen Symmetrielehre das Aufeinanderbezogensein, die gegenseitige Abhängigkeit der Musterelemente, in der sich der Systemcharakter aller Organismen widerspiegelt.
Der Gegensatz zum Kosmos symmetrischer Strukturen ist das Chaos. Hier sind gleich- oder verschiedenartige Elemente, die der Zufall zusammengeführt hat, ohne erkennbare Regelmäßigkeiten im Raum verteilt. Ein ideales Gas, das verschiedene Atome/Moleküle enthält, ist ein perfektes Beispiel für eine solche chaotische Nicht-Struktur.
Die klassischen Symmetrieformen
Die ursprünglich im Bereich der Mineralogie entwickelte Symmetrielehre sah 3 Grundformen der Symmetrie vor: Metamerie, Radiärsymmetrie und Spiegelsymmetrie.
Bei der Metamerie (Longitudinalsymmetrie) sind identische/ähnliche Strukturelemente entlang einer Linie in immer gleicher Orientierung und gleichen Abständen aufgereiht (Metamerie; vgl. Abb. 1 ). Ihre Deckoperation ist die Translation, die Verschiebung von Elementen entlang der Metamerie-Linie. Im einfachsten Fall ist diese Linie eine Gerade, und alle Strukturelemente sind nicht nur gleich gestaltet, sondern auch gleich groß (homonome Segmentierung;Homonomie). Stetige Vergrößerung oder Verkleinerung der Metameren und/oder ihrer Abstände sowie ihre Differenzierung führen zu heteronomer Segmentierung; sie läßt die Verschiebungslinie als strukturbestimmenden Vektor (morphogenetischen Gradienten) erkennen. Bei den Zweigen von Sträuchern und Bäumen ist die sonst meist homonome Metamerie der Sproßachsen (Knoten/Internodien) durch unterschiedlich kräftiges Austreiben von Achselknospen gewöhnlich in heteronome Metamerie verwandelt. Man unterscheidet je nach der Förderung älterer oder jüngerer Achselknospen basitone bzw. akrotone Verzweigungssysteme (Verzweigung; Akrotonie, Basitonie; vgl. Abb. 2 ). Sie entscheiden wesentlich über Wuchsformen und Habitus der Holzgewächse.
Auch bei homonomer Segmentierung kann die Verschiebungslinie als Vektor definiert werden, wenn die Metameren selbst unsymmetrisch sind. Beispiele dafür gibt es vor allem bei Bio-Makromolekülen (Biopolymere) mit ihren unsymmetrischen Monomeren (z.B. L-Aminosäuren, D-Glucose). Polynucleotidstränge (Polynucleotide) besitzen nach der gleichartigen Orientierung der unsymmetrischen (Desoxy-)Ribosylreste ein 3'- und ein 5'-Ende; jede Polypeptidkette (Proteine) besitzt ein Aminoende (Aminoterminus) und ein Carboxylende (Carboxylterminus). In beiden Fällen folgt die Syntheserichtung dem Metamerie-Vektor: DNA-Polymerasen und RNA-Polymerasen verlängern stets nur das 3'-Ende (OH-Ende) von Nucleinsäuresträngen, und der Translationsprozeß (Translation) am Ribosom läuft vom Amino- zum Carboxylterminus durch. Infolge der nicht-periodischen Sequenz (Aminosäuresequenz) von natürlichen Polypeptiden sind auch globuläre Proteinmoleküle (Sphäroproteine) grundsätzlich unsymmetrisch. Durch metamere Aneinanderreihung globulärer Proteineinheiten entstehende Protofilamente (F-Actin, Protofilamente der Mikrotubuli usw.) besitzen daher ebenfalls eine vorgegebene Richtung: Am Plus-Ende ist die Einfügung zusätzlicher Protomeren gegenüber dem Minus-Ende bevorzugt.
Krümmung der Metamerie-Linie in einer Ebene führt zusammen mit heteronomer Segmentierung zu spiraligen Mustern, wie sie auch bei Organismen (z.B. bei den Schalen von Weichtieren; vgl. Abb. 3 ) gelegentlich beobachtet werden können.
Radiärsymmetrie oder Rotationssymmetrie (Strahlensymmetrie, Aktinomorphie) ist gegeben, wenn Symmetrieelemente durch Drehung um eine Symmetrieachse (Achse [Abb.]) zur Deckung gebracht werden können. Deckoperation dieser Symmetrie ist die Rotation. Die Zahl der Symmetrieelemente ist hier – im Gegensatz zu homonomer Metamerie – grundsätzlich begrenzt, die Symmetrieachse ist durch eine definierte „Zähligkeit“ ausgezeichnet. Bei regelmäßigen Vielflächnern (Polyedern), die im Organismenreich vor allem durch Virus-Capside (Viren) und andere Quartärstrukturen von Proteinen exemplifiziert werden, sind in unterschiedlichen Richtungen verlaufende Symmetrieachsen gewöhnlich auch durch verschiedene Zähligkeit ausgezeichnet. Die geläufigsten Beispiele radiärsymmetrischer Biostrukturen werden von Blattwirteln (Wirtel) und Blüten (aktinomorph [Abb.]) geliefert. Auch die Fruchtkörper und Mycelien (Mycel) vieler Pilze sind radiärsymmetrisch. Im Tierreich ist diese Symmetrieform selten; sie beschränkt sich hier vor allem auf sessile oder nur langsam sich bewegende Formen (z.B. Korallen, Thekamöben, Seeigel und Seesterne; vgl. Abb. 4 ) oder auf planktontisch (Plankton [Abb.]) lebende Arten (z.B. Radiolaria [Abb.], Sonnentierchen [Abb.], Quallen).
Die dritte der klassischen Symmetrieformen ist die Bilateralsymmetrie oder Spiegelsymmetrie. Deckoperation der Bilateralsymmetrie ist die Spiegelung, die Zahl der Symmetrieelemente ist 2. Bilateralsymmetrie ist im Tierreich vorherrschend; mehr als 95% der Tierarten zählen zu den Bilateria. Fast stets ist die Spiegelsymmetrie gepaart mit Dorsoventralität (dorsiventral, dorsoventrale Achse), d.h. unterschiedlicher Formung einer Ober- und Unterseite. Dieser komplexen Symmetrie liegen 2 Vektoren zugrunde, die senkrecht aufeinander stehen: Schwerkraft und Bewegungs- (bzw. Wachstums-)Richtung. Spiegelsymmetrie bestimmt auch die Körpergestalt des Menschen sehr weitgehend ( vgl. Abb. 5 ). Für viele Pflanzenfamilien sind „zygomorphe“ Blüten (dorsiventrale Blüte) typisch (Orchideen, Veilchen, Lippenblütler, Rachenblütler [Braunwurzgewächse] usw.). Blattorgane (Blatt) sind fast immer bilateralsymmetrisch. Von vielen Biomolekülen gibt es enantiomorphe (stereoisomere) Formen (Enantiomere [Abb.], Isomerie, optische Aktivität; Kohlenstoff ), von denen im Stoffwechsel gewöhnlich nur eine verwertet wird (z.B. L-Aminosäuren, D-Glucose).
Häufig gibt es bei Organismen Symmetrie-Kombinationen. Beispielsweise sind bei der dispersen (zerstreuten oder wechselständigen) Blattstellung und in vielen Blütenständen (Ähre, Traube) Translation und Rotation kombiniert (Schraubung), bei distich beblätterten Sprossen mit unsymmetrischen Blättern Translation und Spiegelung (Gleitspiegelung: Ulme, Begonie [Begoniaceae]).
Zeitliche Symmetrie, Rhythmen
Wiederholungen von Abläufen entlang der Zeitachse sind als Rhythmen bekannt (Rhythmik, Periodik). Als Symmetrieelement tritt hier eine bestimmte Konstellation in einem System auf, die durch konstante Zeitverschiebung mit der vorangehenden/nachfolgenden zur Deckung gebracht werden kann. Solche zeitliche Metamerien können ohne weiteres als räumliche dargestellt werden (Tier-Fährten, Spinnennetze, Segmentierung als Folge von Entwicklungsrhythmen). Etwa die Hälfte wissenschaftlicher Diagramme haben als Abszisse die Zeitachse. Rhythmen erscheinen dabei als Sinus-Schwingungen oder Überlagerungen (Superposition) von solchen. Viele biologische Vorgänge sind rhythmisch (Biorhythmik), zugleich auf Rhythmen der Umwelt (Tages- und Jahreszeiten, Mondphasen [Lunarperiodizität] und Gezeiten) abgestimmt oder durch sie einreguliert (Chronobiologie [Abb.], Chronopharmakologie).
Eine zeitliche Metamerie von grundlegender Bedeutung für alle Lebewesen ist die Generationenfolge. Die zyklische, in jeder Generation erneut herbeigeführte Rückkehr zu einer einfachsten Ausgangssituation (befruchtete Eizelle, Spore, Brutknospe usw.) führt dazu, daß jedes Entwicklungsstadium (Entwicklung, Embryonalentwicklung) im Fortpflanzungs- (Lebens-)Zyklus zugleich Folge und auch wieder Ursache der Ausgangskonstellation ist. Darauf beruht die (Quasi-) Gleichberechtigung kausaler (warum?) und finaler (wozu?) Betrachtungsweisen in der Biologie (Teleonomie biologischer Systeme; biologischer Zweck, Verursachung, Teleologie – Teleonomie).
Sonderformen der Biosymmetrie
Die Besonderheiten lebender Systeme (Leben) bringen es mit sich, daß es bei ihnen Symmetrie- und Musterformen gibt, die der Mineralogie/Kristallographie fehlen. Wichtig sind Ergänzungssymmetrie sowie stochastische (stochastische Prozesse), funktionale und dynamische Symmetrie. Bei diesen Symmetrieformen wird die Bedingung der Ähnlichkeit von Musterelementen bzw. der Gleichheit ihrer Anordnung teilweise oder ganz aufgegeben. Die Symmetrie äußert sich hier immerhin darin, daß aus dem Vorhandensein von Musterelementen die Existenz und die Orientierung eines oder mehrerer weiterer Elemente postuliert werden können. Dadurch wird der Systemcharakter der Organismen unterstrichen.
Unter Ergänzungssymmetrie („Antisymmetrie“) ist das gesetzmäßige Zugeordnetsein von unähnlichen, aber komplementären Einheiten (Komplementarität) zu verstehen. Diese Form gegenseitiger Zuordnung, die ausnahmslos im Dienste bestimmter Funktionen steht, ist aus der Technik vertraut (Schlüssel/Schloß, Stecker/Dose, Prägestock/Münze, photographisches Positiv/Negativ usw.). Auch bei Organismen stehen antisymmetrische Strukturen häufig im Dienste von Erkennung und/oder Fortpflanzung (Enzym/Substrat, Translokator/Permeand, Rezeptor/Ligand, Antigen/Antikörper). Ergänzungssymmetrisch sind auch die basenkomplementären (Basenkomplementarität) Polynucleotidstränge der DNA-Doppelhelix ( ä Desoxyribonucleinsäuren III ) sowie Codon und Anticodon bei der Translation. Selbstorganisation übermolekularer Aggregate, wie die Bildung von Protein-Quartärstrukturen ( Proteine II ) oder die Formierung von Lipid-Doppelschichten (bimolekulare Lipidschicht, Flüssig-Mosaik-Modell [Abb.]) in Bio-Membranen, beruht ausnahmslos auf Komplementärsymmetrien der molekularen Bausteine. Aus dem makroskopischen Bereich sind der Bau der bei männlichen und weiblichen Tieren korrespondierenden Begattungsorgane (Schlüssel-Schloß-Prinzip) oder die Gelenke der Wirbeltiere zu nennen.
Übermolekulare Biostrukturen werden im allgemeinen zwar ähnlich, aber nicht identisch ausgebildet – man denke an die Blätter eines Baums, an die Schuppen eines Fisches, die Zellen eines Flimmerepithels oder die Mitochondrien einer einzelnen Zelle. Die Schwankungen und Ungleichheiten ergeben sich daraus, daß die Neubildung solcher Musterelemente nicht einem starren Organisationsschema folgt, sondern aus dem Ineinandergreifen von Regulationsprozessen mit entsprechenden statistischen Fluktuationen resultiert. Die fertigen Musterelemente sind in Grenzen variabel: stochastische (statistische) Symmetrie. An die Stelle echter Kongruenz tritt hier die Gleichartigkeit und die morphologische/physiologische Gleichwertigkeit der Musterelemente. Entsprechende Probleme ergeben sich bei der Bildung von Biomustern häufig dadurch, daß Art und Orientierung von Musterelementen nicht genau, sondern nur innerhalb mehr oder weniger großer Streubereiche festgelegt sind. Bekannte Beispiele sind Blattmuster an überwachsenen Mauern, Spaltöffnungsmuster in der unteren Epidermis von Dikotylenblättern, Glomeruli in der Niere oder auch Einzelindividuen in einem Vogel- oder Fischschwarm. Solche Muster sind von präziser Regelmäßigkeit weit entfernt, aber sie sind auch keine Zufallsmuster, schon weil der Abstand zum nächstliegenden Musterelement nicht beliebig variabel ist. Oft kann man alle Übergänge zwischen Mustern von hohem und sehr niederem Ordnungsgrad finden. Umgekehrt ist auch das Erscheinen symmetrischer Muster in vorher ungeordneten Bereichen, d.h. die Entstehung von Ordnung aus Chaos, nicht selten (Beispiele: Entstehung von Blattstellungsmustern an neugebildeten Vegetationspunkten; Bildung von Fruchtkörpern auf einem Pilzmycel; Leitbündelnetze in Blättern, entsprechend Adernnetze in Gliedmaßen). Durch Computersimulation läßt sich zeigen, daß unter einfachen Voraussetzungen (homogene Produktion morphogenetischer Aktivatoren und Inhibitoren mit unterschiedlicher Lebensdauer und Diffusionsrate) regelmäßige Muster entstehen, die an bekannte Biomuster erinnern.
In funktionalen Systemen – alle Organismen sind solche – gibt es eine weitere Form der Symmetrie, die sich überhaupt nicht mehr in Ähnlichkeit oder antisymmetrischer Entsprechung der Systemelemente ausdrückt, sondern in funktionalen Verzahnungen. Solches gilt z.B. für die einzelnen Glieder eines Regelkreises ( Regelung ), die strukturell ganz unähnlich sind, sich aber in ihren Leistungen ergänzen; sie sind kooperativ und wirken synergetisch (Synergetik). Diese funktionale Symmetrie ist morphologisch nicht faßbar. Sie ist eine unanschauliche, aber für lebende (und technische) Systeme besonders typische Form der Symmetrie, die auf Ketten und Netzen funktionaler Antisymmetrien beruht. Je mehr Elemente involviert sind, desto komplexer können die Leistungen eines Systems sein, desto niedriger wird aber zugleich die morphologische Symmetrie. Damit hängt der im allgemeinen niedrige Symmetriegrad von Zellstrukturen (Zelle) zusammen, die jedoch gleichzeitig höchste funktionale Symmetrie aufweisen. Amöben ( vgl. Abb. 6 ) markieren in dieser Hinsicht einen beachtenswerten Extremfall. Daß es nur sehr wenige derart unsymmetrische Organismen gibt, zeigt andererseits, daß die Selektion in Richtung symmetrischer Systeme drückt. Ihre Bildung und ihr Funktionieren erfordert weniger Information als Formierung und Erhaltung unsymmetrischer Individuen.
Lebewesen sind als offene Systeme zwangsläufig auch dynamische Systeme (dynamisches Gleichgewicht), sie können ohne Gestaltveränderungen (Bewegung, Teilung, Wachstum; Evolution) nicht existieren. Diese Grundfunktionen setzen die Möglichkeit von Strukturänderungen voraus, die oft mit Symmetriebrechungen verbunden sind (dynamische Symmetrie). Der Mensch vermag sich trotz der Bilateralsymmetrie seines Körpers ganz anders als nur nach Art eines Hampelmanns zu bewegen. Und ein so hochsymmetrisches Muster wie jenes der Myofilamente (Myofibrillen) in den Sarkomerenquergestreifter Muskulatur ( Muskulatur ) verändert seine Dimensionen drastisch während der Muskelkontraktion (Farbtafel) – allerdings unter Beibehaltung seiner Symmetrien, was mit der Reversibilität dieser Bewegung zusammenhängt. Sind umgekehrt Veränderungen einer hochsymmetrischen Struktur nicht möglich, dann sind auch Lebensvorgänge ausgeschlossen – Virus-Capside belegen dies. Totale Ordnung ist dem Leben sowenig gemäß wie totales Chaos.
Symmetrie und Ästhetik
Durch die Symmetrien der Organismen und auch des menschlichen Körpers, zumal des Gesichts (facial attractiveness), wird der Säugling, der bereits in besonderem Maß zur Gestaltwahrnehmung befähigt ist, in vielfältiger Weise geprägt. Das ermöglicht ihm das frühzeitige Erkennen einer Bezugsperson. Zugleich werden bestimmte Muster und Proportionen gespeichert, die – zusammen mit Variationen im Detail – zeitlebens als schön empfunden werden und schließlich auch zu dem Begriff des „Naturschönen“ hingeleitet haben (Ästhetik). Und obwohl Kunstwerke sicher nicht nur dem Kult des Schönen dienen, sind sie doch häufig (wenn auch unbewußt) so gestaltet, daß sie Symmetrien lebender Systeme widerspiegeln.
Symmetrische Strukturen haben oft ausgesprochenen Signalcharakter. Symmetrische Signale (z.B. Augen, Zähne; Buchstaben und andere Symbole) können durch Kontrastwirkung (Kontrast) vor dem chaotischen Hintergrund-Rauschen einer offenbar überwiegend aus Zufallsmustern aufgebauten Umwelt besonders leicht wahrgenommen werden. Asymmetrie, Chiralität, Disymmetrie, fluktuierende Asymmetrie, Gestalt, Symmetrie-Modell.
Lit.:Brandmüller, J., Claus, R.: Symmetry. Its significance in science and art. In: Interdiscipl. Sci. Rev. 7 (1982), 296. Genz, H.: Symmetrie – Bauplan der Natur. München 1987. Lockwood, E.H., Macmillan, R.H.: Geometric symmetry. Cambridge University Press, Cambridge 1978. Schattscheider, D.: Visions of Symmetry. New York 1990. Sitte, P.: Symmetrien bei Organismen. Biol. in unserer Zeit 14 (1984), 161. Steiner, G.: Spiegelsymmetrie der Tierkörper. In: Naturwiss. Rundschau 32 (1979), 481. Tarassow, L.: Symmetrie, Symmetrie! Heidelberg 1999. Wolf, K.L., Wolff, R.: Symmetrie. Münster 1956.
;)
Symmetrie
Abb. 1:Longitudinalsymmetrie (Metamerie): Laubblatt des Götterbaums (Ailanthus altissima)
;)
Symmetrie
Abb. 2: akrotone (1) und basitone (2) Verzweigung
;)
Symmetrie
Abb. 3: gekrümmte Metamerie-Achse: Schnitt durch ein Nautilus-Gehäuse
;)
Symmetrie
Abb. 4:Radiärsymmetrie: Seestern
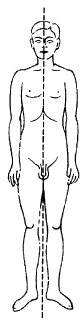
Symmetrie
Abb. 5:Bilateralsymmetrie: Mensch
;)
Symmetrie
Abb. 6:Asymmetrie: Amöbe







Wenn Sie inhaltliche Anmerkungen zu diesem Artikel haben, können Sie die Redaktion per E-Mail informieren. Wir lesen Ihre Zuschrift, bitten jedoch um Verständnis, dass wir nicht jede beantworten können.