Lexikon der Biochemie: Acetylcholin
Acetylcholin, ein sowohl bei Vertrebraten als auch Evertrebraten vorkommender Neurotransmitter in neuromuskulären Synapsen und in den Synapsen folgender Nerven: 1) alle motorischen Nerven zum Skelettmuskel; 2) alle präganglionären Nerven, einschließlich der Nervenversorgung des Nebennierenmarks; 3) alle postganglionären, parasympathischen Nerven; 4) postganglionäre, sympathische Nerven zu den Schweißdrüsen; und 5) einige postganglionäre, sympathische Nerven zu den Blutgefäßen in Skelettmuskeln. Nach der Freisetzung vom Nervenende in die Synapse bindet A. an Rezeptoren des postsynaptischen Neurons und löst damit eine Reaktion aus. Danach wird es vom Rezeptor abgespalten und schnell mit Hilfe der Acetylcholin-Esterase abgebaut (Abb. 1). Nerven, die A. als chemischen Transmitter benutzen, werden cholinerge Nerven genannt.
A. löst – abhängig von seiner Konzentration – zwei verschiedene physiologische Wirkungen aus. Die Injektion kleiner Mengen an A. ruft die gleiche Reaktion hervor wie die Injektion von Muscarin, z.B. einen Abfall des Blutdrucks (wegen Gefäßerweiterung), Verlangsamung des Herzschlags, erhöhte Kontraktion an glatten Muskeln in vielen Organen und starke Sekretion exokriner Drüsen. Dieser muscarine Effekt von Muscarin oder A. wird durch Atropin aufgehoben. Nach Verabreichung von Atropin bewirken größere Mengen an A. eine Erhöhung des Blutdrucks, analog der Wirkung von Nicotin. Nicotinische cholinerge Synapsen werden in neuromuskulären Verbindungen von Vertebraten, bestimmten Ganglien, zentralen Synapsen und den Elektroplaques von Torpedo gefunden. Muscarinische cholinerge Synapsen sind in glatten Muskeln, Herzmuskeln, Ganglien und vielen zentralen Regionen des Gehirns tätig. Im Gehirn und zentralen Nervensystem wird die Zahl der nicotinischen Synapsen von den muscarinischen um das 10-100fache übertroffen. Die nicotinischen cholinergen Rezeptoren werden in Ganglien durch Tetraethylammonium und in neuromuskulären Synapsen durch Curare (Curare-Alkaloide) blockiert. Durch α-Bungarotoxin, einen Bestandteil des Schlangengifts, werden sie irreversibel besetzt. Muscarinische cholinerge Rezeptoren des postganglionären parasympathischen Systems werden durch Atropin und Scopolamin blockiert, die deswegen als parasympatholytische Agenzien bezeichnet werden. Andere Substanzen inhibieren die Aktivität von A.-Esterase und lösen so eine Nervenlähmung aus. Physostigmin z.B. ist ein reversibler Inhibitor, wogegen bestimmte organische Phosphate, die als Insektizide eingesetzt werden, irreversible Inhibitoren sind. Andere Inhibitoren der Acetylcholin-Esterase, wie die organischen Fluorophosphate, zählen zu den wirksamsten chemischen Kampfmitteln (Tab.).
Nicotinische bzw. muscarinische Effekte werden durch nicotinische bzw. muscarinische Rezeptoren vermittelt. Diese Rezeptoren sind die Produkte von zwei verschiedenen Supergenfamilien, und die einzige gemeinsame Eigenschaft dieser Rezeptoren besteht darin, dass sie durch A. aktiviert werden. Der Muscarinrezeptor reagiert langsamer, da er G-Protein-gekoppelte Rezeptoren in seine Antwort mit einbindet. Der vom Effektor ausgelöste Mechanismus hängt vom Subtyp des Rezeptors ab. Er kann z.B. in der Inhibierung von Adenylat-Cyclase, der Bildung von Inositol-1,4,5-triphosphat aus Phosphatidylinositol durch eine spezifische Phospholipase C oder der Modulation (Öffnung) bestimmter K+-Kanäle bestehen [D. Brown Nature 319 (1986) 358-359].
Die muscarinischen Rezeptoren entfalten einen hohen Heterogenitätsgrad, die Primärsequenz hat jedoch bei allen Formen Ähnlichkeit mit der des β2-Adrenorezeptors und des Rhodopsins. Dies lässt vermuten, dass eine gemeinsame Struktur zugrunde liegt, die variiert wird:
1) es sind sieben Transmembran-Segmente vorhanden; 2) der N-terminalen Region fehlt eine Signalsequenz und sie enthält zwei N-Glycosylierungsstellen; 3) die C-terminale Region enthält mehrere Threonin- und Serinreste (Phosphorylierungsstellen); und 4) alle Mitglieder dieser Gruppe sind Zellmembranproteine, die mit G-Proteinen wechselwirken.
Es wurden konventionelle Klonierungsstrategien eingesetzt, mit: a) Reinigung des Rezeptors aus der Großhirnrinde durch Affinitätschromatographie; b) Aminosäuresequenzierung des gereinigten Rezeptors; c) Aufbau von Oligonucleotidsonden auf der Grundlage partieller Aminosäuresequenzen; d) Verwendung dieser Sonden, um cDNA-Bibliotheken zu sreenen. Zusätzlich wurde eine Oligonucleotidsonde für einen Bereich konstruiert, der eine Sequenzhomologie zwischen der zweiten Transmembrandomäne des m1-Rezeptors von Ratten und dem β2-adrenergischen Rezeptor von Hamstern aufweist. Diese Sonde wurde dazu verwendet, cDNA-Bibliotheken aus Rattencortex zu screenen. So wurden fünf pharmakologisch unterschiedliche muscarinische Rezeptorsubtypen (m1-m5) der Großhirnrinde von Ratten und Schweinen cloniert und sequenziert. Durch den Einsatz der Sequenzhomologiesonde wurden weitere Hybridisierungsbanden in Nucleotidbruchstücken der DNA aus Ratten und Menschen gefunden, was auf eine mögliche Existenz weiterer Rezeptorsubtypen hinweist [T. Kubo et al. Nature323 (1986) 411-426; D.C. Hulme et al. Annu. Rev. Pharmacol. Toxicol.30 (1990) 633-673].
m1, m3 und m5 sind mit der Produktion von Inositol-1,4,5-triphosphat gekoppelt, m2 und m4 mit der Inhibierung der Adenylat-Cyclase und der Modulation von K+-Kanälen. Sie werden alle durch Muscarin und Oxotremorin aktiviert und durch Atropin blockiert. Sie weisen außerdem beträchtliche Sequenzhomologie auf und die Strukturanalyse der Sequenz jedes einzelnen Rezeptors weist auf die Existenz von sieben Transmembran-Domänen hin, die die Acetylcholin-Bindungsstelle enthalten.
Die Besetzung des nicotinischen Acetylcholinrezeptors (durch A.) löst eine schnelle Reaktion durch direkte Aktivierung kationenselektiver Ionenkanäle aus (1-2ms) und hat eine Depolarisierung der postsynaptischen Membran zur Folge. Der nicotinische cholinerge Rezeptor wurde aus Elektroplaques von Torpedo californica (Zitterrochen) und Electrophorus electricus (Zitteraal) und aus Muskeln von Vertrebraten isoliert (Abb. 2). In allen drei Geweben handelt es sich um ein einziges Membranprotein (Mr 250 kDa), das aus vier Glycoprotein-Untereinheiten besteht – Mr α 40 kDa (50,1kDa), β 50 kDa (53,7kDa), γ 60 kDa (56,3kDa) und δ 65kDa (57,6kDa) (der erste Wert wurde mit SDS-Gelelektrophorese bestimmt, der Wert in Klammern ist die exakte Mr, die aus der Aminosäure-Zusammensetzung errechnet wurde) – im Verhältnis 2 : 1 : 1 : 1, mit einer durchschnittlich 40 %igen Übereinstimmung der Aminosäuresequenz zwischen allen vier Ketten [B.M. Conti-Troconi et al. Science218 (1982) 1.227-1.229]. Die DNA aller vier Untereinheiten wurde kloniert und sequenziert [M. Noda et al. Nature301 (1983) 251-254]. Verbindungen zur Bestimmung der kovalenten Affinität – [3H]-Bromacetylcholin und 4-(N-maleimido)-3H-benzyltrimethylammoniumjodid – markieren die α-Untereinheit durch Reaktion mit den Cysteinresten 192 und 193. Das bedeutet, dass jede der zwei α-Untereinheiten eine Acetylcholin-Bindungsstelle trägt, also jeder oligomere Rezeptor 2 Bindungsstellen besitzt. Wenn der nicotinische Rezeptor in Liposomen oder planare Lipiddoppelschichten inkorporiert wird, ermöglicht er einen 22Na+-Fluss, der durch A. gefördert und durch α-Bungarotoxin blockiert wird. Der nicotinische cholinerge Rezeptor gehört zu der Gruppe der gesteuerten (engl. gated) Ionenkanäle (hierzu gehören auch die Rezeptoren für γ-Aminobuttersäure, Glycin und 5-Hydroxytryptamin).
Nicotinische Rezeptoren vertebrater neuromuskulärer Knotenpunkte und elektrischer Organe der Fische gehören dem gleichen Subtyp an, den sogenannten peripheren Rezeptoren. Nicotinische Rezeptoren befinden sich außerdem in autonomen Ganglien und im Zentralnervensystem, wo sie unter dem Begriff neuronale Rezeptoren zusammengefasst werden. Letztere sind ebenfalls gesteuerte Kationenkanäle, die aber in struktureller Hinsicht eine heterogene Gruppe bilden und weniger gut charakterisiert sind als die peripheren Rezeptoren.
Alle vier Untereinheiten der peripheren nicotinischen Rezeptoren aus Torpedo oder anderen Quellen weisen ein hohes Maß an Übereinstimmung in der Aminosäuresequenz auf. Besonders enge Homologien bestehen zwischen den α- und β-Peptiden und zwischen den γ- und δ-Peptiden. Dies lässt darauf schließen, dass alle vier Peptide ihren Ursprung in einem einzigen Urgen haben, aus dem sich im Lauf der Evolution Gene für die Ketten des αβ- und des γδ-Typs entwickelt haben. Die Hydrophobizitätsanalyse sagt die Existenz von vier transmembranen α-helicalen Segmenten voraus (M1-M4). Das hydrophile N-terminale Ende jeder Untereinheit enthält Consensus-Sequenzen für die N-Glycosylierung und besteht aus mindestens 200 Resten. Es ist vermutlich extrazellulär angeordnet und enthält eine Cys-Cys-Loop-Domäne, wie sie von anderen gesteuerten Ionenkanälen bekannt ist (Cys-X-X-X-X-X-X-hydrophobisch-Pro-hydrophobisch-Asp-X-X-X-Cys). Da das Protein noch nicht kristallisiert wurde, ist keine Röntgenstruktur-Analyse möglich. Aber mit Hilfe der Elektronenstrahl-Mikroskopie von negativ geladenen Präparaten und Elektronenbild-Analysen von schnell gefrorenen Rezeptorproteinen wurde ein Strukturmodell entwickelt. Im Elektronenbild hat der Rezeptor senkrecht zur Ebene der Membran die Form einer Rosette mit einem Durchmesser von 8nm, die aus fünf Elektronendichtepeaks besteht, die rund um eine zentrale Öffnung angeordnet sind. Diese Rosette bildet das Ende eines Zylinders, der die Membran durchspannt und größtenteils aus der extrazellulären Oberfläche herausragt, z.B. in den synaptischen Spalt. [E.S. Deneris et al. Trends Pharmacol. Sci. 12 (1991) 34-40]
Die Autoimmunkrankheit Myasthenia gravis ist auf das Vorhandensein von zirkulierenden Antikörpern zurückzuführen, die gegen den peripheren nicotinischen Rezeptor gerichtet sind. Die Bindung dieser Antikörper an den Rezeptor hat einen erhöhten Abbau des Rezeptors und eine erniedrigte Effizienz der neuromuskulären Transmission zur Folge. Der Zustand kann durch die Verabreichung von Acetylcholin-Esterase-Inhibitoren teilweise gebessert werden. [J. Newsom-Davis et al. in Clinical Aspects of Immunology, P.J. Lachmann et al. (Hrsg.), Blackwell Scientific Publications, Oxford 1993, 2.091-2.113]
Zur Identifizierung cholinerger Synapsen wird die Acetylcholin-Esterase histochemisch lokalisiert. Dieses Vorgehen basiert auf dem Verfahren von Koelle und Friedenwald [G.B. Koelle Handb. Exp. Pharmakol. 15 (1963) 187-298]. Als Substrat wird Acetyl- oder Butylthiochinolin verwendet und das Produkt – Thiochinolin – wird durch Fällung mit Blei- oder Kupfersalz sichtbar gemacht. Ein spezifischerer Marker für cholinerge Neuronen (Acetylcholin-Esterase ist in dopaminergen Zellen der Substantia nigra ebenfalls vorhanden) ist die Cholin-Acetylase.
A. ist phylogenetisch ein sehr altes Hormon, das auch in Protisten vorkommt. Es ist möglicherweise ein evolutionärer Vorläufer der Neurohormone. In Pflanzen, wie z.B. in den Härchen der Brennessel, befinden sich ebenfalls Spuren von A. Verwandte Verbindungen, z.B. Murexin in den Drüsen bestimmter Gastropoden, sind vermutlich Gifte. [D.S. Mc Gehee u. L.W. Role "Physiological Diversity of Nicotinic Acetylcholine Receptors Expressed by Vertrebrate Neurons" Annu. Rev. Physiol. 57 (1995) 521-546; D.R. Groebe u. S.N. Abramson "Lophotoxin is a Slow Binding Irreversible Inhibitor of Nicotinic Acetylcholine Receptors" J. Biol. Chem. 270 (1995) 281-286; C. Czajkowski u. A. Karlin "Structure of the Nicotinic Receptor Acetylcholine-binding Site" J. Biol. Chem. 270 (1995) 3.160-3.164]
;)
Acetylcholin.Abb. 1. Synthese (im Nervenende) und Abbau (in der Synapse) von Acetylcholin.:
;)
Acetylcholin.Abb. 2. A. Schematische Darstellung der α-Untereinheit des nicotinischen Acetylcholinrezeptors aus Torpedo. M1, M2, M3 und M4 stellen die transmembranen helicalen Regionen dar, die jeweils aus mindestens 20 hydrophoben Aminosäuren, d. h. 51/2 α-Helixwindungen, bestehen. ![]()
sind Kohlenhydratreste. C ist eine Region von M2 in den Untereinheiten β und δ, die den Kanalblocker Chlorpromazin irreversibel binden (Ser254 und Leu257 der β-Untereinheit; Ser262 der δ-Untereinheit). Die Cys-Cys-Schleife in der ausgedehnten N-terminalen extrazellulären Region ist für alle ligandengesteuerten Ionenkanäle charakteristisch. Die benachbarten Cys-Reste (-Cys192-Cys193-) kommen nur in den α-Untereinheiten vor.
B. Anordnung der Untereinheiten des nicotinischen Acetylcholinrezeptors, senkrecht zur Membranoberfläche betrachtet. Die Bindungsstellen für Acetylcholin und α-Toxin liegen auf den α-Untereinheiten.
Acetylcholin.Tab. Wirkstoffe, die cholinerge Systeme beeinflussen.
* Wirkung hauptsächlich auf periphere Ganglien.
Strukturformeln, die in dieser Tabelle nicht aufgeführt sind, können unter einem eigenen Stichwort gefunden werden.
| |
| Muscarinische Antagonisten: Atropin, Scopolamin, Benztropin (blockiert auch die Dopaminaufnahme), Quinuclidinylbromid, Pirenzipin. | |
| Nicotinische Agonisten: Acetylcholin, Nicotin*, Carbachol, Arecolin, Suberyldicholin, Tetramethylammonium*, Phenyltrimethylammonium*, Dimethylphenylpiperazin*. | |
| Nicotinische Antagonisten: D-Tubocurarin, Succinylcholin (Depolarisierung, Desensitivierung), Gallamin, Pempidin*, Mecamylamin*, Hexamethionium*, Pentolinium*, Pancuronium, α-Bungarotoxin. | |
| Inhibitor der Acetylcholinsynthese: 4-Naphthylvinylpyridin. | |
| Pumpeninhibitoren (verhindern das Eintreten von Cholin in die Nervenzelle und führen zum Zusammenbruch der Acetylcholinsynthese): Triethylcholin, Hemicholinium. | |
| Cholin-Esterase-Inhibitoren (werden eingesetzt, um die Cholin-Esterase bei der histochemischen Bestimmung von Acetylcholin-Esterase auszuschalten): Diisopropylphosphofluoridat, Neostigmin, Physostigmin, Edrophonium. | |
| Freisetzungsinhibitor: Botulinus-Toxin. | |
| spezifisch bindende Agenzien: α-Bungarotoxin, Propylbenzilylcholin-Senfgas, Quinuclidinylbenzilat. | |
Carbamylcholin (Carbachol) (stimuliert zuerst Skelettmuskeln, blockiert dann die neuromuskuläre Übertragung). | |
Tetraethylammonium | |
Pentamethonium | |
Hexamethonium | |
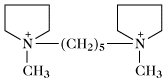 Pentolinium | |
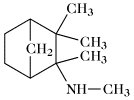 Mecamylamin | |
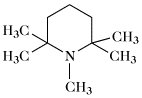 Pempidin | |
 Gallamin (blockiert die neuromuskuläre Übertragung ohne vorhergehende Stimulation). | |
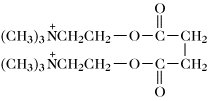 Succinylcholin (Suxamethonium; stimuliert zuerst Skelettmuskeln, blockiert dann die neuromuskuläre Übertragung). | |
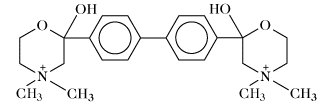 Hemicholinium (verhindert das Eintreten von Cholin in die Nervenzelle). | |
Triethylcholin(verhindert das Eintreten von Cholin in die Nervenzelle). | |
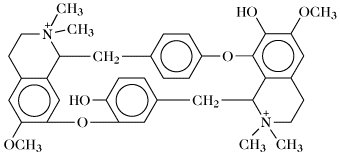 Tubocurarin (blockiert die neuromuskuläre Übertragung ohne vorhergehende Stimulation). | |
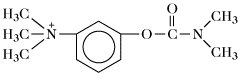 Neostigmin | |
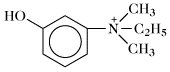 Edrophonium. |







Wenn Sie inhaltliche Anmerkungen zu diesem Artikel haben, können Sie die Redaktion per E-Mail informieren. Wir lesen Ihre Zuschrift, bitten jedoch um Verständnis, dass wir nicht jede beantworten können.