Lexikon der Biologie: Geschichte der Neurobiologie
ESSAY
Robert-Benjamin Illing
Geschichte der Neurobiologie
Einführung
Mit dem Beginn des 19. Jahrhunderts ( vgl. Infobox 1 ) erkannten Karl Friedrich Burdach (1776–1847), Jean-Baptiste-Antoine-Pierre de Lamarck (1744–1829) und Gottfried Reinhold Treviranus (1776–1837) unabhängig voneinander die Notwendigkeit, das Lehrgebäude der Naturgeschichte neu zu gliedern. Unter dem Namen Biologie entstand eine eigenständige Wissenschaft. Die 1839 von Theodor Ambrose Hubert Schwann (1810–1882) und Matthias Jacob Schleiden (1804–1881) begründete Zelltheorie gab der Botanik und Zoologie eine gemeinsame Basis. In seinem Bemühen, diese Theorie zu belegen, führte Schwann zahlreiche Gewebetypen (Gewebe) des tierischen Organismus an, darunter auch das Nervengewebe, vor allem das Rückenmark. Das Nervengewebe wurde als spezifischer Gegenstand biologischer Forschung etwa um die Mitte des 19. Jahrhunderts erkennbar, als sich begründete Zweifel erhoben, ob die Zelltheorie auch für diesen Gewebetyp allgemeine Geltung habe.
Die Erforschung der Strukturen und Funktion des Nervensystems und seiner Beiträge zur Koordination der Körperfunktionen, zu den Wahrnehmungen und dem Verhalten der Organismen wurde als Neurobiologie bekannt. Im Verlauf ihrer Geschichte erhielt die neurobiologische Forschung vielfältige Impulse von Wissenschaften wie der Physik (Biophysik), der Molekularbiologie, der Neurologie und der Psychologie, später auch von der Psychiatrie und den Computerwissenschaften (Computer, Computersimulation). Das gemeinsame Interesse dieser verschiedenen Wissenschaftsbereiche an dem Verständnis von Nerven- und Hirnfunktionen führte sie unter dem Dachbegriff Neurowissenschaft (en) zusammen. Dieser interdisziplinäre Ansatz führte ab 1967 zur Gründung der ersten neurobiologischen Institute, die dem "Unternehmen" eine organisatorische Grundlage gaben und seine weitere Entwicklung förderten.
In dem im 20. Jahrhundert weltweit stetig expandierenden Gebiet der Neurobiologie wurden die verschiedensten Aspekte der neuronalen Strukturen und Funktion von Ionenströmen und neuroaktiven Genen, zellulären Interaktionen, Wahrnehmungsleistungen sowie von Bewußtseinsprozessen (Bewußtsein) thematisiert. Jede der spezifischen Fragestellungen wurde an Modellorganismen untersucht, deren Wahl die Entwicklung der Neurobiologie nachhaltig prägte. Unter den wichtigsten Modellorganismen der Neurobiologie sind zu nennen: der Fadenwurm Caenorhabditis elegans, die Meeresschnecke Aplysia (Seehasen), der Tintenfisch Loligo spec. (Kalmare), die Taufliege Drosophila melanogaster, der Krallenfrosch Xenopus laevis (Krallenfrösche, Xenopus-Oocyten-Expressionssystem), das Huhn (Haushuhn), die Maus (Hausmaus), die Ratte, die Hauskatze (Katzen), der Rhesusaffe und der Mensch.
Die Entdeckung des Neurons
Neuron-Doktrin
Ob sich die Zelltheorie von Schwann und Schleiden auch auf das Nervengewebe übertragen ließ, war zunächst keineswegs sicher. Mit Hilfe des Mikroskops fand man im Nervengewebe zwar viele Zellkörper (Zelle), aber gleichzeitig auch eine große Menge von Faserstrukturen. Robert Remak (1815–1865) kam 1844 nach Untersuchungen an Quetschpräparaten des Rückenmarks zu dem Ergebnis, daß jede Nervenfaser zu einem neuronalen Zellkörper führt. Otto Deiters (1834–1863) war der erste, der aufgrund seiner histologischen Präparate eine klare Unterscheidung zwischen den morphologisch verschiedenartigen Fortsätzen der Neurone (Nervenzelle) treffen konnte ( vgl. Abb. 1 ). Er unterschied den Achsenzylinder, später kurz Axon genannt, von den Protoplasmafortsätzen, die später Faserbäumchen oder, einem Vorschlag des Anatomen Wilhelm His (1831–1904) folgend, Dendriten genannt wurden.
Auf der Grundlage der noch ungenauen histologischen Beobachtungen (Histologie) über das Nervengewebe als Netzwerk von Zellkörpern und Nervenfasern und dem naheliegenden Gedanken eines nervösen Signalflusses entlang der Fasern, beschrieben Joseph von Gerlach (1820–1896) und Camillo Golgi (1843–1926) das Nervengewebe als ein Reticulum, in dem die Zellkörper durch ihre Fortsätze kontinuierlich untereinander verbunden sind. Bei seinen histologischen Untersuchungen am Nervengewebe gelang Golgi die Entwicklung einer Färbemethode, in deren Verlauf einige Zellkörper und Fortsätze von Neuronen und Gliazellen bis in feine Verästelungen hinein schwarz imprägniert wurden. Seine "Schwarze Reaktion", heute als Golgi-Färbung bekannt, lieferte für ein ganzes Jahrhundert detailreiche Einsichten in die Textur des Nervengewebes.
Auch Santiago Ramón y Cajal (1852–1934) bediente sich der Golgi-Färbung, war mit den Interpretationen seiner Kollegen jedoch nicht einverstanden. Er beobachtete, daß sich in einer gegebenen Region des Gehirns immer wieder ähnliche Zellgestalten anfärbten. Die Regelmäßigkeit und Abgeschlossenheit dieser Formen wäre kaum verständlich gewesen, wären die Nervenzellkörper durch ein Netzwerk kontinuierlicher Fasern verbunden. Außerdem waren an den Enden imprägnierter Axone oft besondere Verdickungen ausgebildet, die sog. Endfüßchen. Auf der Grundlage dieser Beobachtungen vertrat Ramón y Cajal schließlich die Auffassung, daß jede Nervenzelle ein Individuum ist, das präzise definierte Grenzen besitzt. Heinrich Wilhelm Gottfried von Waldeyer-Hartz (1836–1921) formulierte 1891 das Prinzip der Nervenzelle als der elementaren Einheit des Nervengewebes und prägte die Bezeichnung Neuron ( vgl. Infobox 2 ). Die Formulierung und Bestätigung der Neuronentheorie war nicht der Beginn, sondern das erste einschneidende Ergebnis neurobiologischer Forschung. Johannes Evangelista von Purkinje (1787–1869) war einer der ersten Forscher, die erkannten, daß die Zellgestalten im Nervensystem keineswegs alle gleichartig sind. In Gewebepräparaten des Kleinhirns sah er, daß die heute als Purkinje-Zellen bekannten Gebilde asymmetrisch und in einer bestimmten Richtung orientiert waren und daß der geschichtete Aufbau des Hirngewebes auf der unterschiedlichen Packung verschiedenartiger Zellkörper beruhte ( vgl. Abb. 2 ).
Ramón y Cajal hat Tausende von Golgi-gefärbten Präparaten des Nervengewebes von verschiedenen Organismentypen und aus den verschiedensten Hirnregionen angefertigt. Es gelang ihm, aus der Betrachtung der statischen Strukturen, die er mit Hilfe des Mikroskops entdeckte, wesentliche dynamische Eigenschaften der Neurone zu erschließen. Sein Prinzip der dynamischen Polarisierung sagt aus, daß die Reizleitung innerhalb einer Nervenzelle vom Empfängerorgan, den Dendriten und dem Zellkörper (Perikaryon), entlang dem Axon zum Senderorgan, der präsynaptischen Endigung, unidirektional verläuft ( vgl. Abb. 3 ). Damit waren den verschiedenen Kompartimenten des Neurons erstmals spezifische Funktionen zugewiesen worden. Wenig später wurde das Prinzip der dynamischen Polarisierung von Charles Scott Sherrington (1857–1952) aufgegriffen. Er argumentierte 1906, daß die Aufgabe des Nervensystems vor allem die Signalintegration (Signal) sei. Es gelang ihm, dieses Konzept auf der Basis des Studiums von Reflexen zu untermauern. Reflexe bilden koordinierte und zweckvolle Verhaltensweisen infolge eines spezifischen Eingangssignals und galten deshalb als Beispiel prototypischen Verhaltens. Durch die Arbeiten von Sherrington wurde die Überzeugung genährt, daß alle zentralen Neurone den Motoneuronen gleichen und sich die integrativen Leistungen des Nervensystems an den Eigenschaften einzelner Neurone studieren lassen.
Aktionspotential
Noch zu Beginn des 19. Jahrhunderts war das Prinzip der Signalleitung (Erregung, Erregungsleitung) im Nervengewebe unverstanden. Luigi Galvani (1737–1798) hatte Froschschenkelpräparate mit Hilfe elektrischer Reizungen zum Zucken (Muskelzuckung) gebracht, ist den Nachweis der Elektrizität (Bioelektrizität) als nervösem Wirkprinzip aber schuldig geblieben. Tatsächlich konnte dieser Nachweis erst erbracht werden, als es gelang, auch kleine Stromflüsse zu messen. Carlo Matteucci (1811–1868) leitete solche Ströme erstmals 1838 am Muskel (Muskulatur) ab. Emil Du Bois-Reymond (1818–1896) beschrieb 1843 einen Nervenstrom, der sich zwischen der Außenfläche und dem Querschnitt eines Nerven ableiten ließ und den er Längs-Querschnitts- oder Ruhestrom (Ruhepotential) nannte. Der Nachweis einer Veränderung dieses Ruhestroms beim Durchgang von Nervenimpulsen, später Aktionspotential genannt, gelang ihm 1849 auch nach chemischer Reizung ( vgl. Infobox 3 ). Damit war der Nachweis erbracht, daß die Nerven nicht nur passive Elektrizitätsleiter sind, sondern selbst elektrische Ströme erzeugen.
Edgar D. Adrian (1889–1977) fand das Aktionspotential in Neuronen aller der von ihm untersuchten Hirnregionen. Es erwies sich überall als großes Alles-oder-Nichts-Signal (Alles-oder-Nichts-Gesetz), das sich ohne Abschwächung vom Zellkörper bis zu den axonalen Endigungen in selbstregenerativer Weise fortpflanzt. Darin zeigte sich das Aktionspotential über die Nervensysteme verschiedener Organismenarten und die verschiedensten Neuronentypen hinweg als universelle neuronale Signalform. Mit Hilfe der von ihnen entwickelten Technik der Spannungsklemme (voltage-clamp-Methode), einer elektronischen Rückkopplungsschaltung, beobachteten Kenneth S. Cole (*1900) und H.J. Curtis am Riesenaxon (Kolossalfasern) des Tintenfisches, daß sich die Leitfähigkeit der Nervenzell-Membran für Ionen während des Aktionspotentials kurzfristig erhöhte. Andrew F. Huxley (*1917) und Bernhard Katz (*1911) fanden mit der von Cole und Curtis entwickelten Methode, daß sich die Größe des Aktionspotentials vermindern ließ, wenn das Axon von einem natriumarmen Medium umgeben war. Dies war die erste einer Reihe von Beobachtungen, die Alan L. Hodgkin (1914–1998), Huxley und Katz in den frühen 1950er Jahren zu einer vollständigen Beschreibung der Ionenströme führte, welche während eines Aktionspotentials über die Nervenzellmembran flossen ( vgl. Abb. 4 ). In ihren Untersuchungen nutzten sie die gerade verfügbar gewordenen Radio-Isotope von Natrium und Kalium, um ionenspezifische Ströme zu messen. Man durfte die lückenlose Beschreibung des für ein Verständnis der Funktionsweise von Nervensystemen jeder Art grundlegenden Signals als eine weitere bahnbrechende Errungenschaft neurobiologischer Forschung feiern. John C. Eccles (1903–1997) erkannte 1957 im Initialsegment der Axone von Motoneuronen die Entscheidungsinstanz, welche über alle erregenden und hemmenden Eingänge integriert und bei Erreichen eines Schwellenwerts die Generierung eines Aktionspotentials (Generatorpotential) initiierte. Wieder schien es, als könnten die Prozesse am einzelnen Neuron als Modell für die Aktionen eines ganzen Gehirns betrachtet werden.
Synapse
Als man den Signalfluß entlang der Axone verstanden hatte, war die Signalweitergabe von einem auf das nächste Neuron zunächst ein noch ungeklärtes Problem. Im Jahre 1897 hatte Sherrington der Kontaktstelle zwischen den Nervenzellen den Namen Synapse gegeben. Das von Ramón y Cajal und Sherrington eingeführte Konzept synaptischer Kontakte war weitestgehend unbestritten, auch wenn es erst der Anwendung des Elektronenmikroskops durch Sanford Palay (*1918) und George Emil Palade (*1912) bedurfte, um die Vorstellungen von der strukturellen Autonomie von Neuronen als zelluläre Individuen im Jahre 1954 endgültig zu bestätigen.
Um das Jahr 1905 applizierte John Newport Langley (1852–1925) mit Hilfe eines feinen Pinsels Nicotin auf isolierte Froschmuskeln. Er stellte fest, daß das Nicotin eine Muskelkontraktion auslöst, wenn es an die neuromuskuläre Synapse gebracht wird; an anderen Stellen der Muskelfasern zeigte Nicotin keinen Effekt. Langley schloß daraus, daß Nicotin auf die postsynaptische Oberfläche der Muskelfaser einwirkt und dort mit einer rezeptiven Substanz reagiert. Otto Loewi (1873–1961) gelang 1920 der experimentelle Nachweis, daß gereizte Nerven tatsächlich eine Substanz freisetzen, welche für die Nervenwirkung verantwortlich ist. Nach den Forschungen von Henry Hallet Dale (1875–1968) handelt es sich dabei um einen Ester des Cholins, das Acetylcholin. Dale hatte noch vermutet, daß jedes Neuron an allen seinen präsynaptischen Endigungen ein und denselben Typ von Transmittermolekülen benutzt. Inzwischen hat sich die Anzahl der bekannten Neurotransmitter vervielfacht, darunter Aminosäuren, Amine, Peptide sowie zyklische Nucleotide und Monoxide (Stickstoffmonoxid), die in jedem Neuron einen spezifischen Cocktail bilden und sich zum Teil sogar von der postsynaptischen zur präsynaptischen Zelle bewegen.
Ricardo Miledi (*1932) erkannte die entscheidende Bedeutung der Bewegung von Calcium-Ionen über die Zellmembran hinweg für die elektro-sekretorische Kopplung an der Synapse. Calcium war seit den Untersuchungen von Sydney Ringer (1835–1910; Ringer-Lösung) als essentieller Bestandteil des Muskel- und Nervengewebes bekannt. Miledi fand, daß infolge der Ankunft eines Aktionspotentials an der präsynaptischen Nervenendigung Calcium aus dem extrazellulären Medium durch spezifische Kanäle in die Zelle hinein strömt. Bleibt dieser Calcium-Strom aus, bleibt die elektrische Aktivität der präsynaptischen Zelle für das postsynaptische Neuron wirkungslos. Miledi beobachtete dann, daß er die Transmitterfreisetzung durch eine Injektion von Calcium in die präsynaptische Endigung in Gang setzen konnte, ohne daß das präsynaptische Neuron elektrisch aktiv war.
Die These einer chemischen Übertragung des Nervenimpulses von Nerven- auf Muskelzellen und zwischen Neuronen wurde von den Neurobiologen zunächst nur zögernd angenommen. Zwei grundlegende Beobachtungen lösten diese Vorbehalte jedoch auf. Katz konnte zeigen, daß Nervenendigungen Signalsubstanzen (Neurotransmitter) in Abhängigkeit von der elektrischen Aktivität paketweise freigeben, und John Heuser und Thomas Reese konnten 1977 mit einer Kombination elektrophysiologischer und elektronenmikroskopischer Methoden den Prozeß einer Verschmelzung präsynaptischer Vesikel mit der Zellmembran bei Aktivität der betreffenden Neurone direkt sichtbar machen. Diese Fortschritte verhalfen der Vorstellung einer Quantelung der Transmitterfreisetzung durch die Exocytose von synaptischen, transmitterhaltigen Vesikeln zu allgemeiner Akzeptanz.
Zwischen 1930 und 1950 wurde eine lebhafte Kontroverse darüber geführt, ob die Signalübertragung an der Synapse elektrisch oder chemisch erfolgt. Während die Befunde an den neuromuskulären Kontaktstellen für eine chemische Übertragung sprachen, schienen die Geschwindigkeit der Übertragung und das Fehlen der peripher nachweisbaren Signalsubstanzen im Zentralnervensystem lange für eine elektrische Übertragung zwischen Neuronen zu sprechen. Die Technik der intrazellulären Messung neuronaler Membranpotentiale revolutionierte das Verständnis der synaptischen Signalübertragung und schien schon endgültig zugunsten der chemischen Hypothese auch für die Synapsen des Zentralnervensystems zu sprechen. Doch gerade auf diesem Stand des Wissens wurde von E.J. Furshpan und D.D. Potter 1957 der Nachweis geführt, daß es einen Typus von Synapsen gibt, heute als gap-junctions bekannt, an denen die Übertragung tatsächlich elektrisch erfolgt.
Heute sind etwa 1000 Proteine der präsynaptischen Endigung bekannt, von denen mindestens 100 unmittelbar an der Regulation der Transmitterfreisetzung beteiligt sind. Die spezifische Interaktion dieser Proteine läßt molekulare Signalnetzwerke für 4 verschiedene Funktionsbereiche erkennen: die Vesikelfusion, die Interpretation der intrazellulären Calcium-Konzentration, der molekulare Gerüstbau innerhalb der präsynaptischen Endigung und die transsynaptische Organisation.
Kanäle und Rezeptoren
Trotz ihrer tiefgreifenden Bedeutung ließ die Analyse des Aktionspotentials durch Hodgkin, Huxley und Katz einen wesentlichen Aspekt des Verständnisses offen, nämlich die Frage nach dem Mechanismus, der es der Nervenzellmembran ermöglicht, die Leitfähigkeit für verschiedene Ionenarten schnell und selektiv zu verändern. Unter Verwendung verschiedener Pharmaka fand Bertil Hille Ende der 1960er Jahre, daß die Natrium- und Kaliumströme durch unterschiedliche Membranporen fließen. Clay Armstrong konnte erste physikalische Eigenschaften dieser Poren definieren, die später u.a. durch Röntgenstrukturanalyse vervollständigt wurden ( vgl. Abb. 5 ). Es bedurfte aber einer Weiterentwicklung der Technik, die 1976 von Erwin Neher (*1944) und Bert Sakmann (*1942) in Form der patch-clamp-Methode vorgestellt wurde, um diese Poren als Kanalproteine (Membranproteine) zu identifizieren, ihre Vielfalt zu erkennen und ihre Eigenschaften genau zu beschreiben ( vgl. Abb. 6 ).
Ein bemerkenswerter Zusammenhang von zunächst beziehungslosen Phänomenen wurde erkennbar, als Katz und Paul Fatt zeigen konnten, daß die Freisetzung von Transmitterstoffen an der postsynaptischen Membran Ionenkanäle öffnen konnte. Dies erinnerte an die Mechanismen, die dem Aktionspotential zugrunde liegen, mit dem Unterschied, daß im Falle der Synapse die Kanäle nicht physikalisch (durch Spannungsänderungen), sondern chemisch (durch Bindung von Transmittermolekülen) geöffnet werden. Dies führte die Befunde von Langley und Loewi mit denen von Hodgkin und Huxley zusammen. Infolge der Einwirkung von Acetylcholin öffnet sich ein neuer Typ eines Ionenkanals, der sowohl für Natrium als auch für Kalium durchlässig ist (Acetylcholinrezeptor, nicotinische Acetylcholinrezeptor-Kanäle, muscarinischer Acetylcholinrezeptor). An inhibitorischen Synapsen (hemmende Synapse) werden dagegen die Aminosäuren GABA (γ-Aminobuttersäure; GABA-Rezeptor-Kanäle) oder Glycin (Glycinrezeptor-Kanäle) freigesetzt, die wiederum andere postsynaptische Kanäle öffnen, welche für Chlorid (Chloridkanäle) oder Kaliumionen (Kaliumkanäle) durchlässig sind. Ein überraschender Befund der modernen Neurobiologie bestand darin, daß spannungsabhängige Kanäle (spannungsgesteuerte Ionenkanäle) molekulare Verwandte von Transmitterrezeptoren sind, d.h., beide sind nach dem gleichen molekularen Motiv konstruiert.
In den 1970er Jahren begannen sich die Hinweise zu verdichten, daß nicht alle Transmitterrezeptoren unmittelbare Wirkung auf die Ionenströme über die Nervenzellmembran haben. Paul Greengard (*1925) bemerkte, daß Transmittermoleküle neben einer schnellen Wirkung an ligandenabhängigen Ionenkanälen (sog. ionotrope Rezeptoren;Liganden-gesteuerte Ionenkanäle) auch langsamere Wirkungen an anderen Rezeptoren erzielen können. Diese langsamen Wirkungen beruhen nicht auf Ionenströmen entlang von Konzentrations-Gradienten (Konzentration), sondern auf Signalkaskaden (Signaltransduktion) über intrazelluläre Moleküle. Deshalb wurden solche Rezeptoren auch metabotrop genannt. Heute ist bekannt, daß über metabotrope Rezeptoren vielfältige Prozesse von einer spät einsetzenden Veränderung der Membranpermeabilität bis zu Modifikation der Genexpression des betreffenden Neurons ausgelöst werden können. Die molekulare Klonierung von Rezeptormolekülen hat gezeigt, daß der Aufbau vieler metabotroper Rezeptoren gleichartige molekulare Motive aufweist: sie haben 7 Membrandurchgänge und entfalten ihre Wirkung über G-Proteine auf Effektormoleküle (Effektor), die auch als sekundäre Botenstoffe (second messengers; sekundäre Boten) bezeichnet werden. Auch das Rhodopsin wurde als Mitglied dieser Molekülfamilie erkannt. Eine andere Gruppe metabotroper Rezeptoren wird von den Rezeptor-Tyrosin-Kinasen (Trk-Rezeptoren) gebildet. Man begann zu verstehen, daß die schnellen synaptischen Aktionen dem Routineverhalten des Organismus zugrunde liegen, die langsamen synaptischen Aktionen dagegen modulatorisch auf die neuronalen Schaltkreise wirken und die Intensität, Dauer und Art des Verhaltens regulieren können.
Wachstumskegel und Dornen
Die morphologische Beschreibung eines Neurons war mit dem Verweis auf Zellkörper, Dendriten und Axon noch nicht vollständig. Nach der Betrachtung histologischer Präparate, die Ramón y Cajal aus embryologischem Gewebe gewann, beschrieb er 1890 spezialisierte Endstrukturen an den Fortsätzen embryonaler Neurone und nannte sie "cones d'accroissement". Er vermutete hinter diesen Wachstumskegeln dynamische Strukturen. Daß es sich dabei tatsächlich um Zellorgane handelt, die sich unter ständiger protoplasmatischer Bewegung langsam vorantasten, erkannte Ross G. Harrison (1870–1959) im Jahr 1908 in ersten Zellkulturexperimenten, bei denen er ein Stück Neuralrohr einer Kaulquappe in einem Tropfen klarer Lymphflüssigkeit (Lymphe) auf einem Glasplättchen unter dem Mikroskop wachsen ließ ( vgl. Abb. 7 ). Mit diesen Experimenten wurde auch der ontogenetische Ursprung (Embryonalentwicklung) der Nervenfasern geklärt, denn Harrison konnte direkt beobachten, daß sie aus Nervenzellkörpern herauswachsen und nicht, wie andere vermuteten, von Gliazellen produziert werden. Mit diesen Beobachtungen begann erkennbar zu werden, daß das Neuron nicht nur den grundlegenden strukturellen Baustein des Nervengewebes darstellt, sondern auch die grundlegende funktionelle Einheit in der Ontogenese des Nervensystems (Nervensystem [Kleindruck]) bildet.
An den Dendriten von Purkinje-Zellen entdeckte Ramón y Cajal 1888 kleine, an der Grenze lichtmikroskopischer Auflösung liegende Strukturen, die sog. Dornen ( vgl. Abb. 8 ). Er vermutete bereits, daß es sich bei diesen Gebilden um Kontaktstellen zwischen Axonen und Dendriten handelt. Die Bestätigung dafür gelang E.G. Gray 1959 mit Hilfe des Elektronenmikroskops. Dornen wurden an den Dendriten verschiedener Typen von Neuronen entdeckt, u.a. an den Pyramidenzellen der Großhirnrinde. Daß die Bedeutung der Dornen über eine einfache Kontaktvorrichtung hinaus geht, wurde deutlich, als M. Colonnier 1968 an ihnen fast ausschließlich erregende Synapsen fand; andere, vor allem inhibitorische Synapsen fand Gray am Schaft der Dendriten sitzen: die Nervensignale, die Neurone über Dornen erreichen, stellen folglich eine spezifische Auswahl dar.
E. Fifkova wies 1982 darauf hin, daß sowohl Wachstumskegel als auch Dornen ein Cytoskelett (Zellskelett) aufweisen, das ausschließlich aus Actin besteht, während die anderen Teile des Neurons außerdem Mikrotubuli und Neurofilamente als typische Cytoskelettbestandteile enthalten. Da der Wachstumskegel als bewegliche Struktur bereits bekannt war, wurde vermutet, daß auch Dornen bewegliche Strukturen sein könnten. Die Kombination selektiver Eingänge und die durch die Organisation des Cytoskeletts nahegelegte Beweglichkeit führte zur Hypothese, daß Neurone die Gewichtung ihrer Eingänge durch Veränderung der Dornengestalten gezielt verstellen können. Tatsächlich wurden seit 1969 systematische Veränderungen der Gestalt der Dornen in Zusammenhang mit Verhaltensvariationen beobachtet, von denen man später lernte, daß sie innerhalb von Sekunden stattfinden können.
Neuronale Zellverbände
Regionale Differenzierungen des Nervengewebes
Die Neuro-Embryologie wurde durch W. His begründet. Er studierte die anatomischen und zellulären Formen des Neuralrohrs während der Embryonalentwicklung des Menschen ( Embryonalentwicklung I ) und entwickelte Vorstellungen, nach denen die Neurone nicht nur zelluläre Einheiten, sondern auch nutritive, funktionelle und genetische Einheiten sind und sowohl als Empfänger als auch als Sender von Signalen agieren, die das ontogenetische Wachstum des Nervensystems steuern.
Im Jahr 1915 entdeckte Calvin B. Bridges (1889–1938) eine Mutante (Entwicklungsmutante) der Taufliege Drosophila, bei der die Struktur eines Körpersegments auf ein anderes Körpersegment übertragen war: das die Halteren tragende Segment war in dieser bithorax (Bithorax-Komplex) genannten Mutante durch ein zweites Segment mit einem Flügelpaar ersetzt worden (homöotische Mutante [Abb.]). Die Wirkung dieser Mutation hatte sich offenbar auf ein Körpersegment beschränkt. Edward B. Lewis (*1918) untersuchte weitere Gene mit der Eigenschaft, die Identität ganzer Körpersegmente zu transformieren, und fand, daß solche sog. homöotischen Gene einer Colinearitätsregel folgend auf dem Chromosom in derselben Reihenfolge angeordnet sind wie die Körpersegmente, auf die sie wirken. Ein Verständnis der molekularen Wirkungsweise homöotischer Gene gelang erst, nachdem man gelernt hatte, sie zu klonen (Klonierung). Walter J. Gehring (*1939) fand, daß alle homöotischen Gene von Drosophila einen Abschnitt enthielten, über den sie sich kreuzweise hybridisieren (differentielle Hybridisierung) ließen. Dies war die Entdeckung der Homöobox, ein etwa 180 Basenpaare langes Stück DNA, das für ein 60 Aminosäuren langes Protein codiert (Homöobox-Gene), welches seinerseits eine hohe Bindungsaffinität an spezifische Abschnitte anderer Gene zeigt. Als man 1984 versuchte, Hybridisierungen auch mit Genen des Hühnchens, der Maus und des Menschen durchzuführen, zeigte sich zwischen den Genen dieser grundlegend verschiedenen Organismen eine erstaunliche Homologie, die auch die Colinearitätsregel mit einschloß. Damit wurde die Frage akut, ob auch die so identifizierten Gene der Wirbeltiere zu einer Segmentidentität beitragen. Im Gegensatz zum Nervensystem von Drosophila sind Gehirn und Rückenmark Höherer Wirbeltiere nicht offenkundig segmentiert organisiert, die vorübergehende Bildung von Rhombomeren während der Ontogenese des Rhombencephalons (Rautenhirn) hat jedoch die Vermutung geweckt, daß sich dahinter eine segmentale Organisation verbirgt. Robb Krumlauf konnte 1989 zeigen, daß jedes der 8 Rhombomeren im Hinterhirn (Metencephalon) der Maus durch ein einzigartiges Muster der Expression von Homöobox-haltigen Genen ausgezeichnet war ( vgl. Abb. 9 ). Diese Beobachtungen zeigen in eindrucksvoller Weise, daß es Netzwerke genetischer Elemente gibt, welche die Koordination komplexer Musterbildungen (Musterbildungsgene) übernehmen und offenkundig über weite Zeiträume der Evolution konserviert geblieben sind.
Vor allem die Augenentwicklung ( Induktion ) ist zu einem wichtigen Modell der Neurogenetik geworden. Die eindrucksvollsten Beispiele sind bisher die Gene eyeless und Pax-6. Als Gehring mehrere ektopische Fehlexpressionen (ektopische Expression) von eyless in Drosophila induzierte, kam es zur Bildung ektopischer Augen. Obwohl das genetische Wirkgefüge, das zur Bildung des völlig andersartig gebauten Wirbeltierauges beiträgt, noch weitgehend unverstanden ist, fand man durch Sequenzvergleiche von Genen, daß dabei viele der Gene eine Rolle spielen, die auch bei der Entwicklung des Komplexauges der Insekten von Bedeutung sind. Durch diese Befunde wurde ein bis dahin für ausgeschlossen erachteter monophyletischer Ursprung der verschiedensten Augentypen im Tierreich denkbar. Auch im Vorderhirn der Wirbeltiere wurden während der Ontogenese spezifische Muster von Genexpressionen entdeckt. Für seine Differenzierung werden z.B. die Gene Gsh1 und Gsh2 aktiviert; entsprechend führte der Verlust der Genfunktion von Gsh in Knockout-Mäusen zu einer Unterbrechung der Vorderhirn-Differenzierung. Die Funktion dieser Gene besteht in der Regulation von Zelladhäsionsmolekülen (Adhäsine), unter denen sich die Cadherine befinden. Lokale Genexpression und spezifische Regulation von Zelladhäsionsmolekülen tragen zur Regionalisierung des Nervengewebes bei und führen zur Bildung und Aufrechterhaltung von embryonalen Abschnitten, Hirnkernen, Hirnrindenschichten (Hirnrinde) und neuronalen Schaltkreisen.
Entwicklung neuronaler Verknüpfungen
Zur Beantwortung der zentralen Frage der Neuroembryologie, wie nämlich die komplexen Muster der Verbindungen im Nervensystem zustande kommen, gab es zunächst nur Spekulationen. Zur Erklärung dieses Phänomens bedienten sich Ramón y Cajal und einige seiner Zeitgenossen Begriffen wie Chemotaxis, Chemotropismus, Galvanotaxis und Neurotropismus. Paul A. Weiss (1898–1989) vermutete 1924, daß das Nervensystem zunächst als diffuses Netzwerk, eine Art "tabula rasa", gebildet wird, in dem erst durch Erfahrung und Übung des Individuums ungeeignete Verbindungen nach einem Resonanzprinzip eliminiert werden, um schließlich ein funktionell angepaßtes Kommunikationssystem hervorzubringen. Roger W. Sperry (1913–1994) konnte jedoch zeigen, daß die Entstehung axonaler Verbindungen keineswegs orientierungslos erfolgt. Er nutzte die Regenerationsfähigkeit (Regeneration) retinaler Ganglienzellen von Fischen und Amphibien aus, die ein normales Sehvermögen einige Tage gach Durchtrennung des optischen Nerven (Opticus) wieder ermöglicht. Drehte er das Auge vor dem Regenerationsprozeß um 180°, erwiesen sich nach erfolgter Regeneration auch die Verhaltensreaktionen um 180° fehlgeleitet ( vgl. Abb. 10 ). Diese Situation blieb für die Dauer des Lebens der Tiere unverändert, wurde also durch Lernprozesse (Lernen) nicht korrigiert. Sperry schloß aus diesen Befunden, daß die Neurone ihren Zielpunkt im Gehirn unabhängig von ihrer retinalen Position ansteuern. Die Innervation der spezifischen Zielgebiete im Mittelhirn erfolgte auch nach erzwungener Umleitung des Wachstumsweges. Es wurde erkennbar, daß präzise Verhaltensleistungen direkt auf die spezifische Verbindung zwischen Neuronen angewiesen sind.
Die Erklärung dieser Phänomene fand ihren Niederschlag in Sperrys Chemoaffinitätshypothese, bei deren Formulierung im Jahr 1963 er sich auch auf Vorarbeiten von J.N. Langley stützte. Nach dieser Hypothese erwerben Zellen in jeder neuronalen Population während der frühen Ontogenese einen bestimmten Satz chemischer Marker, die ihre Position innerhalb der Population spezifizieren und dazu genutzt werden, ein spezifisches räumliches Muster auch in den Terminationsgebieten der von dieser Population ausgehenden Nervenbahnen auszubilden. Lange wurde die Bestätigung der Chemoaffinitätshypothese durch das Fehlen von Nachweisen für die dafür erforderlichen chemischen Marker behindert. Alternative Theorien kamen unter der Bezeichnung Morphogenetik auf, die versuchten, das geordnete axonale Wachstum (Axogenese, axon guidance, Axonwachstum) durch mechanische Interaktionen zwischen den auswachsenden Axonen zu erklären, dabei jedoch nur in begrenztem Maße erfolgreich waren. Erst Anfang der 1980er Jahre wurde man bei der Suche nach chemischen Markern fündig, die Konzentrationsgradienten von Wegweisermoleküle aufbauen: die Familie der Eph-Rezeptoren erfüllte die Kriterien, die nach der Chemoaffinitätshypothese von Positionsmarkern gefordert wurden.
Zelltod und Neurotrophine
Die intrinsischen Fähigkeiten, regionale Differenzierungen auszubilden und spezifische axonale Verbindungen herzustellen, sind jedoch nicht die einzige gestaltende Kraft bei der Entstehung eines voll funktionsfähigen Nervensystems. Vorstellungen von "Fermenten" oder "stimulierenden Substanzen", die infolge einer Nervenverletzung gebildet werden, um Regenerationsprozesse zu fördern, wurden bereits von Ramón y Cajal und J.F. Tello entwickelt. Sie konnten jedoch noch keine experimentelle Evidenz zugunsten dieser Hypothese aufweisen. Viktor Hamburger (1900–2001) beobachtete Anfang der 1930er Jahre, daß die einem bestimmten Flügel zugeordneten Motoneurone im Hühnchen-Embryo verschwinden, wenn die Flügelknospe zuvor experimentell entfernt wurde. Unter dem Einfluß seines Lehrers Hans Spemann (1869–1941) interpretierte Hamburger das Verschwinden dieser Zellen dadurch, daß ihnen das passende induktive Signal (Induktionsfaktor, Induktionsstoff) aus der Zielregion ihrer Axone fehlte. Rita Levi-Montalcini (*1909) bestätigte diese Befunde und fand eine korrespondierende Degeneration von Neuronen im sensorischen Ganglion, das den Flügel, dessen Entwicklung durch Ablation verhindert wurde, normalerweise versorgt. Tatsächlich beeinflußte die Entfernung der Flügelknospe keineswegs die Proliferation, Wanderung oder Differenzierung der Neurone, sondern wirkt sich erst in einem sekundären Prozeß auf das Überleben der Neurone aus. Die genaue Beobachtung dieser Vorgänge zeigte 1949, daß ein bestimmter Anteil der Neuronenpopulation im sensorischen Ganglion auch im Normalfall degeneriert. Diese erste Dokumentation des programmierten Zelltods (Apoptose, Paraptose) fand zunächst wenig Beachtung. Erst im Laufe der 1970er Jahre häufte sich die Evidenz, daß der selektive Zelltod nicht nur in allen untersuchten Systemen des Zentralnervensystems vorkommt, sondern auch den beträchtlichen Anteil von 30–70% der ursprünglich bereitgestellten Neuronenpopulation erfaßt. Durch Untersuchungen an Caenorhabditis elegans fand Robert Horvitz, daß unter einem Dutzend Genen, die in der einen oder anderen Weise mit der Apoptose verbunden sind, die Gene ced-3, ced-4 und ced-9 eine Schlüsselrolle spielen ( vgl. Abb. 11 ). Während einige dieser Gene für Caspasen codieren, die Proteine angreifen, welche für die Lebensfähigkeit der Zelle wichtig sind, wirken die Produkte der anderen Gene solchen Aktionen entgegen. Die in dem Fadenwurm entdeckten Sequenzen der Apoptose-Gene decken sich in erstaunlichem Maße mit homologen Genen des Wirbeltier-Genoms.
Hamburgers Befunde schienen 1934 anzudeuten, daß die sich entwickelnden peripheren Strukturen ein spezifisches Agens produzieren, das über den retrograden axonalen Transport in die spinalen Motoneurone gelangt und dort die Population der Motoneurone quantitativ reguliert. Elmer Buecker beobachtete 1948, daß bestimmte Typen von Sarkomen (Krebs) von sensorischen, aber nicht von motorischen Axonen innerviert werden ( vgl. Abb. 12 ). Die diesen Prozessen zugrundeliegende Substanz wurde 1956 von Stanley Cohen (*1922) und Rita Levi-Montalcini aus dem Schlangengift gereinigt und Nervenwachstumsfaktor (Nerve growth factor, NGF) genannt. Sie fanden, daß der Nervenwachstumsfaktor das Überleben, das Auswachsen von Neuriten und die Neurotransmitterproduktion eines bestimmten Nervenzelltyps fördert, nämlich der Neurone des Sympathikus im peripheren Nervensystem. Erst 30 Jahre später wurde durch Y. Barde und Hans Thoenen ein zweites Neurotrophin isoliert, dessen Wirksamkeit diesmal für Neurone des zentralen Nervensystems festgestellt wurde: der brain derived neurotrophic factor (BDNF). Heute sind 7 Neurotrophine (neurotrophe Faktoren) bekannt, die Entdeckung weiterer neurotropher Substanzen ist zu erwarten. Ihre Funktion läßt sich inzwischen nicht mehr auf die Förderung des Wachstums von Neuronen begrenzen. Je nach den spezifischen Umständen können Neurotrophine auch als Neuromodulatoren wirken (z.B. in Zusammenhang mit der Langzeitpotenzierung), sie können Ströme an spannungsabhängigen Ionenkanälen beeinflussen, und sie können auch das Absterben von Neuronen bewirken.
Neuronale Netzwerke
Der Beginn von Versuchen, Funktionen von Nervenzellverbänden aus einem theoretischen Ansatz heraus zu verstehen, begann mit den Arbeiten von Warren McCulloch (1898–1969) und Walter Pitts im Jahre 1943. Sie konnten zeigen, daß selbst einfache neuronale Netzwerke im Prinzip jede Art von arithmetischer und logischer Operation ausführen können ( vgl. Abb. 13 und vgl. Abb. 14 ). Dieser Ansatz wurde von Norbert Wiener (1894–1964) und John von Neumann (1903–1957) aufgegriffen, die vorschlugen, Computer zu konstruieren, deren Architektur dem Aufbau natürlicher Gehirne entlehnt war. Donald O. Hebb (1904–1985) erweiterte den Gedanken logisch operierender Neurone um den Aspekt des Lernens. Er formulierte 1949 eine hypothetische Regel (Hebbsche Regel), nach der die Modifikation der Übertragungseigenschaften einzelner Synapsen in Abhängigkeit von der zeitlichen Ordnung prä- und postsynaptischer Aktivität vonstatten gehen könnte ( vgl. Infobox 4 ). Hebbs energischer, spekulativer Schritt einer neuronalen Formulierung von Lernprozessen hat Psychologen, Neurobiologen und Computerwissenschaftler gleichermaßen elektrisiert und die Entwicklung von Maschinen initiiert, die von der Funktionsweise des Nervensystems inspiriert wurden. Die aus diesen Ansätzen entstandenen neuronalen Netzwerke sind Maschinen aus künstlichen Neuronen. Das Äquivalent eines Computerprogramms enthalten sie in den spezifischen Regeln, nach denen ihre Elemente untereinander kommunizieren. Frank Rosenblatt stellte 1958 mit seinem Mark 1 Perceptron den ersten erfolgreichen "Neurocomputer" vor, gleichzeitig entwickelte Karl Steinbuch (*1917) die sog. Lernmatrix. Während der 1960er Jahre kam es zu einem erbitterten Wettstreit zwischen den Schulen der neuronalen Netzwerke und der künstlichen Intelligenz um finanzielle Ressourcen, den die erstere aus Mangel an zündenden Ideen zu verlieren drohte ( vgl. Infobox 5 ). John Hopfield gelang es jedoch, die Idee der Neurocomputer wieder zu beleben, so daß es Mitte der 1980er Jahre zu einer Explosion von Forschungsergebnissen kam, die in zunehmendem Maße dazu beitrugen, zentrale Funktionsaspekte natürlicher Nervensysteme und Gehirne aus einer neuen Perspektive heraus zu verstehen. Teuvo Kohonen (*1934) entwarf Netzwerke, in denen sich konkrete oder abstrakte Merkmale im Lauf von Lernprozessen durch Veränderung der interneuronalen Kontakte nach Art einer Landkarte systematisch sortieren. Stephen Grossberg (*1939) erfand Netzwerke, die in Anlehnung an die Architektur des Isocortex hochgradig rückgekoppelt waren und sich bei der Modellierung von Wahrnehmungsprozessen bewährt haben. D.E. Rumelhard und G.E. Hinton haben 1985 neuronale Netzwerke entwickelt, die als Backpropagation-Modelle bekannt wurden und aus mehreren Schichten künstlicher Neurone bestehen. Auf der Grundlage solcher Backpropagation-Modelle haben neuronale Netze erstaunliche Leistungen erzielen können, z.B. bei der Erkennung von Gesichtern (Gesichtererkennung) oder beim Erlernen des Vorlesens von Schriftsprache (Sprache).
Neuronales System und Wahrnehmung
Lokalisationslehre
Nach Jahrzehnten spekulativer Zuordnungen psychologischer Leistungen zu besonderen Hirnregionen wurde die Lokalisation spezifischer Funktionen durch Charles Bell (1774–1842) und François Magendie (1783–1855) erstmals experimentell begründet. Sie entdeckten unabhängig voneinander das Gesetz der Rückenmarkswurzeln, das heute als Bell-Magendie-Gesetz bekannt ist. Dieses Gesetz besagt, daß die vom Rückenmark ausgehenden Nervenwurzeln nicht gleichartig sind, sondern daß die hinteren (dorsalen) Wurzeln Wahrnehmungsfunktionen (sensorischen Funktionen), die vorderen (ventralen) Wurzeln jedoch Bewegungsfunktionen (motorischen Funktionen) dienen. In Übereinstimmung mit dieser Erkenntnis formulierte Johannes Peter Müller (1801–1858) im Jahr 1826 das Gesetz der spezifischen Sinnesenergie. Danach ist die "Sprache" der Nervenzellen universell: eine spezifische Wahrnehmung kommt nicht durch eine besondere Art von Nervenaktivität zustande, sondern dadurch, daß die Nervenzellen an einem bestimmten Ort des Gehirns aktiv werden. Marie Jean Pierre Flourens (1794–1867) legte das Hinterhirn eines lebenden Hundes frei und beschrieb, wie er durch mechanische Reizung des Gewebes an der Hirnoberfläche Muskelkontraktionen hervorrufen konnte. Als er diese mechanischen Reize vom Rückenmark beginnend immer weiter vorne ansetzte, kam er an einen Punkt, von dem an keine Muskelkontraktionen mehr auslösbar waren oder, wenn er von vorne beginnend rückwärts ging, wieder einsetzten. Der Punkt des Übergangs lag reproduzierbar im Bereich des Mittelhirns. Flourens schloß aus dieser Beobachtung, daß der untere Hirnstamm motorisch, das Vorderhirn jedoch sensorisch sein müsse. Eduard Julius Hitzig (1838–1907) und Gustav Theodor Fritsch (1838–1897) reizten die Hirnrinde elektrisch, obwohl diese seit Flourens als unerregbar galt. Vom frontalen Cortex der freiliegenden Hirnoberfläche eines Hundes konnten sie jedoch motorische Reaktionen der dem Reizort gegenüberliegenden Gliedmaßen auslösen. Bei Reduzierung der Reizstärke gelang es sogar, einzelne Muskelgruppen dieser Gliedmaßen zur Kontraktion zu bringen.
Die funktionelle Spezialisierung lokaler Hirnregionen war inzwischen auch den Neurologen auffällig geworden. Paul Broca (1824–1880) bestätigte eine Beobachtung des britischen Landarztes Marc Dax (1770–1837), wonach Patienten mit plötzlichem Sprachverlust (Aphasie) oft Verletzungen einer bestimmten, immer links gelegenen Hirnrindenregion aufweisen. Auch Carl Wernicke (1848–1905) beschrieb aphasische Patienten, deren Sprachunvermögen jedoch von anderer Art war und deren Hirnverletzungen zwar auch links, aber weiter posterior lagen. Wegen ihrer Bedeutung für die Sprachfunktionen wurde die linke Hemisphäre bald die dominante genannt. Erst viel später gelang es, Funktionen zu identifizieren, die von der rechten Hirnhemisphäre dominiert werden (Lateralisation, Asymmetrie).
Anhand der Zellstrukturen gelang es Korbinian Brodmann (1868–1918), 47 corticale Regionen der menschlichen Großhirnrinde (Cytoarchitektonik; Rindenfelder) strukturell zu unterscheiden: die enge Verzahnung von Struktur und Funktion des Gehirns wurde zunehmend offenkundig. Am Gehirn des Menschen erwies sich die Gültigkeit der Lokalisationslehre als besonders dramatisch. Um die Mitte des 20. Jahrhunderts eröffnete Wilder Graves Penfield (1891–1976) den Schädel und die Hirnhäute von Patienten, deren schwere Epilepsie chirurgisch behandelt werden sollte. Zur präoperativen Orientierung stimulierte Penfield die Hirnrinde der wachen Patienten mit einem schwachen elektrischen Strom an verschiedenen Stellen und ließ sich berichten, ob und was die Patienten dabei erleben. Je nach Reizort konnten das einfache Lichtblitze oder undefinierbare Geräusche sein, manchmal kam es zu Zuckungen von Muskeln. In bestimmten, temporal gelegenen Rindenregionen konnte es dagegen passieren, daß die Patienten von komplexen Erinnerungsbildern berichteten. Heute liegen Karten der funktionellen Differenzierung der Hirnrinde von eindrucksvoller Komplexität vor. Allein die Sehrinde des Rhesusaffen umfaßt nach heutiger Kenntnis mehr als 30 unterscheidbare Areae (Area).
Konzepte hierarchischer Hirnorganisation
Paul Emil Flechsig (1847–1929) studierte im späten 19. Jahrhundert den Verlauf der Reifung der menschlichen Hirnrinde und unterschied 3 Typen von Hirnrindenregionen, von denen nur die primären Zonen sensorische Signale aus subcorticalen Regionen erhalten. Die beiden anderen nannte er Assoziationscortex, weil sie zu einer Stufe der Entwicklung reiften, während der die Kinder in der Wahrnehmung die verschiedenen Sinne untereinander und mit Bewegungen zu verbinden lernten. Flechsigs Schema wurde von Psychologen aufgegriffen und im Sinne neuroanatomischer Korrelate psychologischer Leistungen interpretiert.
Rafael Lorente de Nó (1902–1990) machte sich auf die Suche nach elementaren Funktionseinheiten im Gehirn und beschrieb die grundlegende Architektur des zentralen auditorischen Systems und des vestibulären Systems sowie den völlig andersartigen Aufbau der Formatio reticularis. Ausdrücklich wies er auf die grundsätzliche Ähnlichkeit des Isocortex von Maus und Mensch hin. Um herauszufinden, in welchen funktionellen Abhängigkeiten die verschiedenen Hirnregionen untereinander standen, gab es damals nur 2 methodische Ansätze: die detaillierte Analyse von Patienten mit Hirnverletzungen und die Beobachtung des Verhaltens und der cerebralen Degenerationsprozesse nach gezielten Läsionen an Versuchstieren. Paul MacLean (*1914) entwickelte unter Anwendung dieser Methoden die Vorstellung, daß das menschliche Gehirn aus 3 Teilen aufgebaut ist, die 3 Entwicklungsebenen der Evolution entsprechen: das dreieinige Gehirn. In seinem Schema wird die Entwicklungsstufe der Reptilien durch den sog. R-Komplex (zu dem er die Basalganglien rechnete), die des paläo-mammalischen Gehirns durch das limbische System (ein Terminus, den er 1951 prägte) und die der Säuger durch den Iso- (oder Neo-)cortex repräsentiert ( vgl. Abb. 15 ). Jede dieser Stufen hat ein charakteristisches Verhaltensrepertoire hervorgebracht, das unter geeigneten Umständen am Menschen beobachtbar ist. MacLean schrieb dem R-Komplex eine zentrale Rolle in der sozialen Kommunikation und Konformität, dem limbischen System das Reich der Gefühle (Emotionen) und das rationale Denken dem Isocortex zu. Später durchgeführte Studien zur Konnektivität des Gehirns haben zahlreiche strukturelle Aspekte dieses Schemas bestätigen können.
Der Beobachtung von Franz Alexander Nissl (1860–1919) folgend, daß es nach Läsionen der Hirnrinde zu einem Verlust an Nervenzellkörpern im Thalamus kommt, untersuchte W.E. LeGros Clark die thalamo-corticalen Projektionen. Trotz der Limitierung dieser Methode, die darin begründet war, daß eine Verletzung des Axons keineswegs alle betroffenen Nervenzellkörper zugrunde gehen läßt, wurden mit ihr wesentliche Einsichten in die Organisation des Gehirns erarbeitet. Ein weitergehendes Studium der Organisation des Gehirns der Wirbeltiere erwies sich wegen der dichten Lage und engen Verflechtung der vielen Millionen Neuronen zunächst als unmöglich. Erst die Entwicklung gut geeigneter Tracing-Methoden ("Bahnmarkierungen") erbrachte einen gewaltigen Zuwachs an Informationen über die Hirnorganisation. Die erste experimentelle Methode, Faserzüge im Gehirn darzustellen, war die Marchi-Technik (benannt nach Vittorio Marchi) der 1930er Jahre, die 1946 durch Paul Glees verbessert wurde. Den Durchbruch bei den Methoden, axonale Faserzüge jeder Art von ihrem Ursprung bis zu den präsynaptischen Endigungen experimentell sichtbar zu machen, gelang Walle J.H. Nauta (1916–1994) im Jahr 1952. Die Darstellung neuronaler Projektionen mittels des anterograden Transports (axonaler Transport) radioaktiv markierter Aminosäuren wurde 1965 von A.V. Taylor und P. Weiss vorgestellt. Tracer-Techniken (Tracer) auf der Basis von Enzymen kamen ab 1972 zur Anwendung, nachdem es J.H. LaVail und M.M. LaVail gelang, Meerrettich-Peroxidase intraaxonal wandern zu lassen ( vgl. Abb. 16 ). Schließlich wurden von Hans Kuypers (1925–1989) verschiedene Fluorochrome gefunden, die keine chemische Nachbehandlung des Gehirngewebes mehr erforderten und durch kombinierten Einsatz die Beantwortung komplexerer neuroanatomischer Fragestellungen erlaubten. Intrazelluläre Markierungsmethoden (Markierung) wurden entwickelt, die nun die Golgi-Methode in der Darstellung feinster morphologischer Details der Neurone übertraf, und Entwicklungen auf den Gebieten der Histochemie und Immuncytochemie (Immunhistochemie; vgl. Abb. 17 ) eröffneten Möglichkeiten der Lokalisation nahezu beliebiger Moleküle in Schnitten durch das Nervensystem.
Ausgestattet mit einem wachsenden Methodenrepertoire traten die Systematiker der Hirnforschung an. Als einer der Begründer der modernen Neuroanatomie erarbeitete Nauta in genauer Kenntnis neurologischer und psychiatrischer Phänomene grundlegende Einsichten in die strukturelle Organisation funktioneller Systeme des Gehirns, vor allem des präfrontalen Cortex, der Basalganglien und des limbischen Systems. Thomas P.S. Powell (1923–1996) erzielte umfangreiche Befunde zur Organisation intracorticaler und corticothalamischer Verbindungen sowie des olfaktorischen Systems und beschrieb mit Hilfe des Elektronenmikroskops die Grundmuster synaptischer Schaltkreise in den verschiedensten Vorderhirnregionen. Edward G. Jones und Jon H. Kaas legten detaillierte Kartierungen der Großhirnrinde und des Thalamus vor, mit deren Kenntnis die Basis dafür gelegt wurde, ein genaueres Verständnis des integrativen Signalflusses im Vorderhirn zu erlangen.
Rezeptive Felder
Halden K. Hartline (1903–1983) versuchte erstmals im Jahr 1938, die Aktivität von Ganglienzellen in Abhängigkeit von der Belichtung der Retina (Netzhaut) zu registrieren. In seinen Versuchen am Froschauge fand er, daß die Ganglienzellen spontan Aktionspotentiale generieren. Die Frequenz dieser Entladungen konnte moduliert werden, wenn Licht auf eine bestimmte Region der Retina fällt. Die Region auf der Netzhaut, von der aus sich die Aktivität einer Ganglienzelle modulieren läßt, nannte er ihr rezeptives Feld (Kontrast). Horace B. Barlow konnte 1953 zeigen, daß die rezeptiven Felder retinaler Ganglienzellen ein inhibitorisches Umfeld besitzen. Er entwickelte ein Konzept, demzufolge einzelne Neurone als Detektoren ausgebildet sind, welche das Vorliegen von Reizen melden, die für die Wahrnehmung und das Verhalten des betreffenden Organismus bedeutungsvoll sind (feature detectors). Barlow vermutete, daß das Nervensystem in einer Weise gebaut ist, deren Effizienz darin liegt, daß Wahrnehmungsobjekte durch eine möglichst geringe Anzahl von Neuronen detektiert und repräsentiert werden. Aufgrund dieser Neurondoktrin der Wahrnehmung ( vgl. Infobox 6 ) ebnete Barlow den Weg für eine große Anzahl von Forschungsansätzen, die erneut von der Überzeugung getragen wurden, daß sich das Gehirn und die neuronalen Grundlagen der Wahrnehmung durch ein Studium einzelner Neurone erschließen lassen würden. Stephen W. Kuffler (1913–1980) fand, daß sich die Methoden zur Untersuchung rezeptiver Felder auch an Säugern erfolgreich einsetzen ließen. Auf diesem Stand der Forschung begannen David H. Hubel (*1926) und Torsten N. Wiesel (*1924) mit ihrem Forschungsprogramm, das eine eindrucksvolle Sequenz zunehmender Komplexität von rezeptiven Feldern im visuellen System, vor allem in der primären Sehrinde, zu Tage förderte ( vgl. Abb. 18 ).
Die Individualität von Wahrnehmungsobjekten resultiert aus einer spezifischen Kombination von Komponenten, z.B. Form (Formensehen; Bildwahrnehmung), Farbe (Farbensehen), Textur und Bewegung (Bewegungssehen, Bewegungswahrnehmung). Das Erkennen dieser individuellen Objekte (Objektwahrnehmung; Gestaltwahrnehmung) sollte also darauf beruhen, daß die spezifische Kombination von Komponenten wahrgenommen wird. Eine Möglichkeit, dies zu erreichen, ist die Konvergenz der Signale von Neuronen, welche die Gegenwart einzelner Komponenten signalisieren, auf Neurone höherer Ordnung, deren Aktivität dann die Gegenwart einer bestimmten Kombination der Komponenten repräsentiert. Jenseits der eigentlichen Sehrinde wurden tatsächlich Neurone mit hochkomplexen visuellen rezeptiven Feldern entdeckt. Charles Gross fand 1972 im inferotemporalen Cortex (corticales Areal auf dem Temporallappen) eines Affen Neurone, deren rezeptive Felder offenbar so eingerichtet waren, daß die maximale Aktivierung durch die Präsentation der Silhouette einer Hand erreicht wurde. 10 Jahre vergingen, bis dieser zunächst unglaubliche Befund bestätigt und erweitert wurde. In der Folgezeit wurden in naheliegenden Hirnregionen Neurone entdeckt, die spezifisch auf die Konturen eines artspezifischen Gesichts (Gesichtererkennung) reagierten. Wegen ihrer spezifischen Filtereigenschaften wurden solche Neurone Großmutterzellen, gnostische Neurone oder Pontifikalzellen genannt. William T. Newsome konnte zeigen, daß die Spezifität der rezeptiven Felder relevant für das Verhalten des betreffenden Organismus sein kann, indem er die Aktivität solcher Neurone in Wahrnehmungssituationen manipulierte und das Versuchstier daraufhin zu einer motorischen Antwort veranlaßte, die der neuronalen Aktivität und nicht der Wahrnehmung entsprach. Neurone mit hochkomplexen spezifischen Funktionen wurden auch in zentralnervösen Strukturen anderer sensorischer Systeme gefunden. Im Hippocampus entdeckte John O'Keefe Zellen, die immer dann Aktionspotentiale feuerten, wenn sich das Versuchstier innerhalb einer bekannten Umgebung an einer bestimmten räumlichen Stelle befand; benachbarte Neurone waren an anderen Raumpunkten aktiv. O'Keefe nannte die durch diese Neurone konstituierte Abbildung die kognitive Karte (kognitive Landkarte) des Raums.
Die Erklärung der Wahrnehmung komplexer Gegenstände durch die Konvergenz fester axonaler Verbindungen auf höherrangige Neurone muß aber aufgrund der Vielfalt möglicher Wahrnehmungen zu einer kombinatorischen Explosion führen, für die selbst das menschliche Gehirn keine hinreichend große Anzahl von Neuronen bereithält. Als Alternative wurde deshalb von P.M. Milner und Christoph von der Malsburg vorgeschlagen, die Repräsentation spezifischer Wahrnehmungen zusätzlich durch einen verteilten Code zu erklären: individuelle Neurone sollten danach in einem Moment an der Repräsentation eines Wahrnehmungsobjekts, im nächsten Moment aber an der eines anderen teilnehmen können. Wolf Singer (*1943) hat umfangreiche Belege zusammentragen können, daß die Bindung der Neurone zu einem assembly, das zu einem bestimmten Zeitpunkt eine konkrete Wahrnehmung repräsentieren könnte, durch die zeitliche Korrelation der von den gerade zusammengehörigen Neuronen generierten Aktionspotentiale erfolgen könnte. In Übereinstimmung mit dem Konzept von assemblies zeigte Apostolos P. Georgopoulos, daß im motorischen Cortex Richtung und Größe von Willkürbewegungen nicht durch einzelne Neurone, sondern über eine ganze Population von Neuronen hinweg mit Hilfe eines Populationscodes spezifiziert werden.
Plastizität neuronaler Systeme
Nach der Zurückweisung der Lehren von Weiss durch Sperry blieb die Vorstellung einer erfahrungsabhängigen Veränderung der Hirnstruktur lange Zeit außerhalb des Vorstellungsvermögens der Neurobiologen. Erst in den späten 1960er Jahren wurde durch G. Raisman und Gerald Schneider nachgewiesen, daß zumindest Teile des Gehirns plastisch sind. Besonders eindrucksvoll war 1970 der Nachweis durch Hubel und Wiesel, daß der visuelle Cortex sensorische Aktivität braucht, um zu seiner vollen strukturellen und funktionellen Differenzierung zu gelangen. Diese Beobachtungen führten zu einer sprunghaften Zunahme der Erforschung der Plastizität.
Die Forschungsergebnisse, die nun zusammengetragen wurden, schienen zunächst zu belegen, daß die Stimulation durch Umweltreize nur für eine relativ kurze Phase während der frühen postnatalen Ontogenese wichtig und wirksam für die Entwicklung des Nervensystems ist. Diese Phasen nannte man die kritischen Perioden (sensible Phase; Prägung). Die Möglichkeit plastischer Veränderungen der Hirnrinde oder sogar subcorticaler Hirngebiete adulter Säuger erschien noch immer nicht realistisch. Diese Zurückhaltung blieb bestehen, obwohl doch bekannt war, daß sich sensorische und motorische Fähigkeiten zeitlebens durch Übung verbessern lassen. Noch immer war die enge Verflechtung von Hirnfunktion und Hirnstruktur nicht hinreichend fest in den Vorstellungen der Forscher verankert. In dem Moment jedoch, als moderne Tracer-Techniken und Mikroelektrodenableitungen (Ableitung, Mikroelektrode) kombinierbar wurden, begann eine reiche Ernte von Ergebnissen zur Plastizität der Hirnrinde und subcorticaler Strukturen zu allen Lebensaltern der Säuger. Es gab wesentliche Unterschiede in den plastischen Reaktionsmöglichkeiten junger und ausgereifter Gehirne, aber prinzipiell wurde in beiden Fällen das vollständige Repertoire plastischer Reaktionen beobachtet: Zu- oder Abnahme des Volumens von Neuronen und Neuronengruppen, Modifikationen topographischer Karten, Umleitung von Axonen, Auswachsen oder Zurückziehen dendritischer Fortsätze, Zelltod, Mitose und molekulare Veränderungen vielfältiger Art.
Der Gesang der Vögel, der von Spezies zu Spezies sowie zwischen den Geschlechtern unterschiedlich ist, bot den Neurobiologen ein besonders aufschlußreiches Modell, um genetische Faktoren von Lernprozessen zu unterscheiden und die mit Lernvorgängen einhergehenden hirnorganischen Veränderungen zu beobachten. Junge Zebrafinken (Prachtfinken) werden von ihren Eltern aufgezogen, beginnen aber erst als ausgewachsene Männchen mit dem arttypischen Gesang. Man entdeckte, daß die Wahrnehmung des Gesangs der Eltern wesentlich für die Ausbildung des eigenen Gesangs ist. Der Erwerb des Gesangs steht unter hormoneller Kontrolle. Normalerweise singen nur die Männchen der Zebrafinken. Werden sie kastriert, singen sie nicht. Arthur Arnold und Fernando Nottebom gelang es 1980, Weibchen zum Singen zu befähigen, indem ihnen unmittelbar nach dem Schlüpfen geeignete Metabolite des Testosterons injiziert wurden. Seine Wirkung entfaltet das Hormon durch Bindung an Neurone im Hirnstamm, Mittelhirn und Vorderhirn. Die betreffenden Zellen sind normalerweise in den männlichen Gehirnen größer als in den weiblichen. Die Hormonbehandlung (Hormone, Neurohormone) der Weibchen führte erwartungsgemäß zur Vergrößerung der betreffenden Neurone und Hirnregionen.
Nach der Ausbildung eines grundlegenden, genetisch initiierten Bauplans kommt es also nach Maßgabe neuronaler Aktivität zu einer Präzisierung der neuronalen Verbindungen. Die umfassende Bedeutung dieser Entwicklungsstrategie wurde erst durch das Studium an Säugern deutlich. Muster neuronaler Aktivität greifen in 2 Stufen in die Ontogenese des Nervensystems ein. Carla Shatz konnte zeigen, daß schon in pränatalen Entwicklungsphasen intrinsische Aktivitätsmuster generiert werden, die maßgeblich an der Musterbildung neuronaler Verknüpfungen beteiligt sind. Kaspar-Hauser-Versuche haben überzeugende Belege dafür geliefert, daß die spezifischen Muster neuronaler Aktivität, die durch Sinneserfahrung, also durch extrinsische Quellen, indiziert werden, abermals entscheidende Wegweiser für eine umfassende Hirnentwicklung liefern. Colin Blakemore und Michael Stryker konnten zeigen, daß das Ausbleiben spezifischer neuronaler Erregung spezifische synaptische Verbindungen und Eigenschaften rezeptiver Felder einzelner Neurone beeinflussen kann. Bemerkenswert ist dabei, daß die molekularen Signalwege, welche die Bildung synaptischer Verknüpfungen nach Maßgabe neuronaler Aktivitätsmuster vermitteln, teilweise identisch sind mit denen, die im Zuge von Lernprozessen und der Langzeitpotenzierung genutzt werden. Nach heutigem Wissen müssen Gene, sensorische Stimulation, Eigenstimulation und Gedächtnissysteme (Gedächtnis) zusammenspielen, um ein angepaßtes koordiniertes Verhalten herauszubilden. Das relative Gewicht dieser Faktoren kann von Spezies zu Spezies und von System zu System variieren.
Verhalten und Bewußtsein
Vitalismus, Behaviorismus, Funktionalismus
Noch im 19. Jahrhundert war bei anerkannten Wissenschaftlern wie Johannes P. Müller, Gustav Theodor Fechner (1801–1887) und Hans A.E. Driesch (1867–1941) ein Gedankengut lebendig, das außerphysikalische Faktoren, z.B. die Seele (Leib-Seele-Problem) als umfassendes Formprinzip, in die Erklärung neurobiologischer Beobachtungen mit einbezog. Die von Hermann Ludwig Ferdinand von Helmholtz (1821–1894) und Du Bois Reymond geführte "Physikalismus"-Bewegung (Physikalismus) begann jedoch solche "vitalistischen Spekulationen" auszugrenzen (Vitalismus – Mechanismus;Mechanismus, Vitalismus): Erklärungen biologischer Phänomene einschließlich tierischen und menschlichen Verhaltens hatten ausschließlich auf beobachtbare Größen Bezug zu nehmen, das Phänomen Bewußtsein wurde als grundsätzlich unerklärbar ausgewiesen. Dem behavioristischen Ansatz zufolge, den John B. Watson (1878–1958) besonders eloquent propagierte, sollten nur noch das äußerlich sichtbare und meßbare Verhalten der Organismen anerkannt werden: "Die Zeit scheint gekommen zu sein, daß die Psychologie alle Verweise auf ein Bewußtsein verwerfen muss". Der Behaviorismus arbeitete mit Reiz-Reaktions-Schemata als Erklärungsmodell und versuchte den Organismus nach dem Vorbild einer Maschine zu verstehen, ohne dabei wesentliche Unterschiede zwischen Mensch und Tier zu machen. Er betrachtete sie als "black box" (Black-box-Verfahren) und wollte ihre Funktionsweise allein aus den Reizen und den daraus resultierenden Reaktionen erschließen. Von einem eigenständigen Innenleben der Organismen ist nicht die Rede; eine "Vererbbarkeit" von Verhaltensmerkmalen wurde geleugnet. Der Behaviorismus kam jedoch in Erklärungsnöte, als es um ein Verständnis komplexerer Lernvorgänge ging. Vor allem die Art und Weise, wie Menschen Sprache lernen, war mit einem Reiz-Reaktions-Schema nicht zu erklären. Das Ende des Behaviorismus wurde durch die Entwicklung des Computers eingeleitet. Die Entdeckung, daß Computer zu Leistungen gebracht werden konnten, die bis dahin dem Menschen vorbehalten waren, belegte eindrucksvoll, daß intelligentes Verhalten (Intelligenz) sehr wohl ein Innenleben erfordert, dessen genaues Verständnis nun unverzichtbar erschien, um komplexes Verhalten zu erklären. Aus diesen Einsichten entsprang der Funktionalismus, der besagt, daß mentale Zustände nicht nur Nebenprodukte von Reflexbahnen sind, sondern selbst kausal wirksame Zustände darstellen (vor allem vertreten durch Hilary Putnam, Daniel Dennett sowie, in gewissem Sinne, Patricia und Paul Churchland). Technische Systeme wie Computer werden als prinzipiell umfassendes Modell für alle neurobiologischen Funktionen anerkannt. Folgt man diesem Denkansatz, so wird der Mensch eines Tages aus einem Computer einen vollwertigen Organismus konstruieren können, indem er ihn mit einer geeigneten Mechanik versieht und zum Roboter (Bionik, künstliche Intelligenz, künstliches Leben) ergänzt.
Den Einsichten des Funktionalismus verdanken wir, daß Aufmerksamkeit und Bewußtsein wieder als wichtige Konzepte in der Neurobiologie erscheinen. Für viele Neurobiologen und Philosophen bleibt aber fraglich, ob funktionelle Zustände im Sinne des Funktionalismus identisch sein können mit dem subjektiven oder qualitativen Charakter (Qualia) unseres Erlebens.
Schlafen, Wachen und Bewußtsein
Die Möglichkeit, elektrische Aktivität des Nervengewebes mit Hilfe von Ableitungen durch Schädel und Kopfhaut hindurch zu messen (Elektroencephalogramm, EEG), wurde 1875 von Richard Caton (1842–1926) und 1890 von Adolf Beck (1863–1942) entdeckt. Caton registrierte "zarte Ströme variabler Richtung" am Kaninchenkopf, deren Rhythmus sich zwischen dem Zustand des Schlafens (Schlaf) und des Wachens (Vigilanz) unterschied und sich durch sensorische Stimulation deutlich beeinflussen ließ. Während im Schlafzustand hohe Spannungsschwankungen mit geringer Frequenz beobachtbar waren (Alpha-Rhythmus; Alpha-Wellen), führten Wachheit und Aufmerksamkeit zu einer Zunahme von hochfrequenten Spannungsschwankungen mit geringer Amplitude ( vgl. Abb. 19 ). Beobachtungen an Patienten mit chronischer Lethargie und Läsionen im Bereich des Mittelhirns führten zu der Vorstellung eines Schlafzentrums im Hirnstamm. Frédéric Bremer (1892–1959) durchtrennte 1935 im Gehirn eines Säugetieres die Faserzüge zwischen den Colliculi superiores und inferiores (Cerveau isolé; Colliculi) und fand, daß das betreffende Tier dadurch in einen Dauerschlaf fällt. Der normale Schlaf-Wach-Rhythmus (Chronobiologie [Farbtafel], Chronopharmakologie) bleibt jedoch erhalten, wenn die Durchtrennung am caudalen Ende des Hirnstamms (Encéphale isolé) erfolgte. Eine partielle Durchtrennung des Hirnstamms auf Höhe des Mittelhirns unterbrach den normalen Schlaf-Wach-Rhythmus nur dann, wenn auch die medialen Faserzüge im Zentrum des Hirnstamms unterbrochen wurden. Diese Ergebnisse wiesen darauf hin, daß die Verbindungen zwischen der Formatio reticularis und dem Vorderhirn intakt bleiben müssen, um einen normalen Schlaf-Wach-Rhythmus zu ermöglichen. Elektrische Reizung der Formatio reticularis weckte dann auch Versuchstiere aus dem Schlaf. Diese Beobachtungen führten Giuseppe Moruzzi (1910–1986) und Horace Winchell Magoun (1907–1991) zur Theorie eines aufsteigenden reticulären Aktivierungssystems (ARAS), dessen Aktivität den Zustand der Wachheit herbeiführt und das EEG in seine desynchrone Form versetzt. Hans Berger (1873–1941) studierte das EEG in der Überzeugung, daß diese neue Technik auch Aufschluß über Bewußtseinsprozesse des Menschen geben könne.
Besonders deutlich wurde die Desynchronisation des EEG im Zustand der Aufmerksamkeit. Das 1958 von Donald Broadbent vorgestellte Konzept, demzufolge im Zustand selektiver Aufmerksamkeit die Ressourcen des Gehirns auf Teile der sensorischen Stimuli gebündelt werden, schien sich in zunehmendem Maße zu bestätigen. Mit Hilfe moderner bildgebender Verfahren wie der Positronenemissionstomographie (PET) oder funktionellen Kernspintomographie (fMRI) konnte Michael Posner Netzwerke von Neuronen identifizieren, die an der Steuerung der Aufmerksamkeit beteiligt sind. Auch die Wahrnehmung von Objekten bestimmter Form, von Geräuschen mit spezifischer Bedeutung, das Auftauchen von Seelenregungen, wie Angst, Humor und Liebe, oder spezifische Denkleistungen gehen mit neuronalen Aktivitäten an ganz bestimmten Orten des Gehirns einher, sog. neuronale Korrelate des Bewußtseins. Die Liste solcher neuronaler Korrelate bewußten Erlebens unterstreicht die Richtigkeit der Lokalisationslehre – und erweitert sie. Aufgrund der Natur der Daten, die diese Liste enthält, wird sie aber nicht dazu beitragen können zu erklären, welche kausale Beziehung bewußtes Erleben zu hirnorganischen Vorgängen hat und warum sich Schmerz oder Freude überhaupt nach etwas anfühlen.
Lernen und Gedächtnis
Schon seit dem Altertum galt das Erinnerungsvermögen als eine zentrale Eigenschaft des menschlichen Geistes. Hermann Ebbinghaus (1850–1909) begründete die psychologisch-experimentelle Untersuchung des Gedächtnisses noch ohne Bezug auf die Veränderungen zu nehmen, die möglicherweise dabei im Gehirn stattfinden. Richard Semon (1859–1918) verankerte bereits die Gedächtnisfunktionen in biologischen Prozessen und prägte den Terminus Engramm (Gedächtnisspur). Ramón y Cajal, für den die strukturelle Veränderbarkeit neuronaler Zellverbände infolge von Lernprozessen offenkundig war ( vgl. Infobox 7 ), dachte dabei vor allem an amöboide Bewegungen von Gliazellen. Diese Vorstellung wurde aber von Rudolf Albert von Kölliker (1817–1905) kritisiert, der erkannte, daß solche Bewegungen viel zu langsam seien, um Gedächtnisspuren niederlegen zu können. Karl Spencer Lashley (1890–1958) unterstrich später diesen Einwand, verwarf aber gleichzeitig den Gedanken an erfahrungsabhängige Veränderungen von Synapsen als Wunschdenken. Mitte des 20. Jahrhunderts bestand Ratlosigkeit darüber, wie Gedächtnisinhalte im Gehirn niedergelegt werden.
In dieser Situation entstanden 2 grundverschiedene Forschungsansätze: 1) In Anlehnung daran, daß das "genetische Gedächtnis" in Nucleinsäuren codiert ist, wurde nun nach biochemischen Grundlagen (Biochemie, Biochemie [Geschichte der] ) des individuellen Gedächtnisses gesucht. In den 1960er Jahren wurde aus den Gehirnen von Ratten, die mit Elektroschocks darauf trainiert wurden, sich entgegen ihrer Vorliebe für Dunkelheit im Hellen aufzuhalten, ein 6–10 Aminosäuren langes Peptid isoliert, das von seinem Entdecker Georges Ungar (1906–1978) Scotophobin genannt wurde. Die Injektion dieses Peptids in das Peritoneum (Bauchhöhle) untrainierter Ratten sollte zu einem vermehrten Aufenthalt dieser Tiere im Hellen geführt haben und beweisen, daß spezifische Lerninhalte durch spezifische Moleküle repräsentiert werden. Diese Versuche haben zu ihrer Zeit für großes Aufsehen gesorgt, doch blieben Bestätigungen aus. 2) Das von D.O. Hebb formulierte neurophysiologische Postulat des Lernens (s.o.) eröffnete konkrete Ansätze für neurobiologische Experimente. Erste Meldungen Ende der 1970er Jahre, nach denen Hebbsche Synapsen im Nervengewebe entdeckt wurden, gaben den Anstoß zu einer außerordentlich fruchtbaren Entwicklung am Knotenpunkt von Molekularbiologie, Neurobiologie und Psychologie. Das Phänomen der Langzeitpotenzierung, das von Tim Bliss und Terje Lomo erstmals 1973 umfassend beschrieben wurde, manifestierte sich in einer aktivitätsabhängigen Veränderung der synaptischen Interaktion von Neuronen, die an die Vorstellungen von Hebb erinnerten. Vor allem Eric Richard Kandel trug durch seine Arbeiten an Aplysia dazu bei, Schritt für Schritt die molekularen Ereignisse zu entschlüsseln, die der synaptischen Plastizität zugrunde liegen. Die Vorstellung, daß es sich bei der Langzeitpotenzierung wirklich um ein zelluläres Korrelat von Lernprozessen handelt, schien durch die Tatsache gestützt zu werden, daß sich die Langzeitpotenzierung besonders deutlich an Gewebekulturen des Hippocampus zeigen ließ, einer Hirnrindenregion, deren Beteiligung an Lern- und Gedächtnisprozessen schon länger bekannt war. Heute wissen wir, daß Langzeitpotenzierung an Synapsen von Neuronen in den meisten Hirnregionen stattfinden kann und eine zentrale Rolle auch in der ontogenetischen Ausbildung neuronaler Verbindungen und bei plastischen Prozessen spielt.
Verhalten und Kognition
Das Verhalten von Individuen aus allen Höheren Tierstämmen beruht auf der integrativen Aktivität des Nervensystems. Einige der genauesten Untersuchungen zum Verständnis und zur Entwicklung von Verhalten kamen von Ethologen (Ethologie, Ethologie [Geschichte der] ) und Psychologen, die ihre Beobachtungen noch ohne expliziten Bezug auf das Nervensystem dargestellt hatten: die neurobiologischen Einsichten in die Funktionsweise des Nervensystems waren in der 1. Hälfte des 20. Jahrhunderts noch nicht hinreichend detailliert. Die Entdeckung von komplexen Verhaltensweisen frisch geschlüpfter oder neugeborener Organismen, die offenkundig in die Welt, in der sie noch keine Erfahrungen sammeln konnten, hineinpaßten, war ein Phänomen, das sorgfältigen Beobachtern auffallen mußte. Charles Darwin (1809–1882) beobachtete das ausgeklügelte Verhalten von Insekten und erkannte, daß die Bewegungsabläufe in hohem Maße stereotyp (Stereotypie) sind: die Tiere verharrten bei den angeborenen Bewegungsabläufen, auch wenn ihnen durch äußere Eingriffe das Erreichen des Ziels ihrer Aktivitäten erleichtert wurde. Konrad Lorenz (1903–1989) und Nikolaas Tinbergen (1907–1988) erkannten, daß diese Art programmierten Verhaltens keineswegs auf Wirbellose begrenzt, sondern auch unter Wirbeltieren weit verbreitet ist. Tinbergen konnte nachweisen, daß sog. Schlüsselreize ganze Verhaltenssequenzen auslösen können. In einer berühmt gewordenen Serie von Versuchen untersuchte er 1951 das Paarungsverhalten (Balz, Balzkette, Begattung; Signal) eines Fisches, des Stichlings. Der männliche Stichling entwickelt während der Paarungszeit eine rote Bauchseite, welche die Geschlechtsgenossen zu aggressivem Verhalten (Aggressivität, Aggression) provoziert, Weibchen dagegen zu Annäherungsversuchen veranlaßt. Tinbergen fand heraus, daß ein naturgetreues Stichlingsmodell keine solche Reaktionen auszulösen vermag, solange es keine rote Bauchseite hat. Dagegen waren schon sehr grobe Modelle wirksam in der Auslösung des artspezifischen Verhaltens, sobald ihre Unterseite die richtige Farbe aufwies ( vgl. Abb. 20 ). Tinbergen hat diese Ergebnisse noch nicht in neurobiologischen Termini beschrieben und die Existenz spezifischer, jahreszeitlich modifizierbarer rezeptiver Felder in den visuellen Regionen des Fischgehirns noch nicht vermuten können.
Behauptungen, nach denen das Verhalten des Menschen eine angeborene Komponente hat, haben früher heftige Reaktionen ausgelöst. Spätestens seit der Entdeckung durch Jérome Lejeune (†1997) im Jahr 1959, daß das Down-Syndrom, das offenkundig mentale Defizite beinhaltet, auf einem genetischen Defekt beruht, mußte der Einfluß der Gene anerkannt werden. Vergleiche der Lebensschicksale eineiiger Zwillinge (Zwillingsforschung; Adoptionsstudie, Anlage-Umwelt-Diskussion) haben belegen können, daß auch Persönlichkeitsmerkmale und Intelligenz erbliche Komponenten haben. Zweifellos nehmen aber die Plastizität und das Lernen bei der Entwicklung des Menschen im Vergleich zu anderen Säugern den höchsten Stellenwert ein. Eric H. Lenneberg hat mit umfangreichem Material die Tatsache belegt, daß der Erwerb der Muttersprache beim Menschen besondere neurobiologische Dispositionen in Anspruch nimmt, die nur für eine begrenzte Lebenszeit (Lerndisposition, sensible Phase) zur Verfügung stehen. Das Erlernen einer Zweitsprache erfolgt unter Beteiligung anderer, räumlich weiter verteilter Regionen des Gehirns.
Als Primat, der in komplexen sozialen Zusammenhängen lebt (Soziobiologie), ist besonders der Mensch darauf angewiesen, subtile Körper-, Hand- und Augenbewegungen (Bewegung, Hand) und Gesichtsausdrücke (Mimik) hervorzubringen und wahrzunehmen. Die auf diesen Wegen ausgetauschten Botschaften können aufrichtig oder unaufrichtig, freundlich oder unfreundlich, aggressiv oder unterwürfig sein. Ein wesentlicher Teil unseres Erfolges bei der Erschließung von Nahrung, Geschlechtspartnern und Verbündeten beruht auf unserem Geschick, diese Botschaften schnell und genau zu entziffern (Biokommunikation, Kommunikation). Mit dem Aufkommen bildgebender Verfahren gelingt es in zunehmendem Maße, auch solche komplexen kognitiven Leistungen (Kognition) mit der neuronalen Aktivität in konkreten Hirnregionen in Verbindung zu bringen. So zeichnet sich ab, daß das "soziale Gehirn" vor allem aus 3 Regionen besteht: dem Sulcus temporalis superior (Furche im Bereich des Temporallappens), dem orbitofrontalen Cortex (orbitaler Teil des präfrontalen Cortex) und der Amygdala. Menschen mit krankheits- oder verletzungsbedingten Schädigungen dieser Regionen verlieren die Orientierung in ihrem sozialen Verhalten und bei ethischen Wertzuweisungen (Bioethik, Ethik).
Ausblick
Die Geschichte der Neurobiologie lehrt uns, daß die neurobiologische Forschung in ihrem Bemühen, die Funktionen von Neuronen, Neuronenverbänden und Gehirnen auszuloten, in den vergangenen 200 Jahren überaus erfolgreich war. Die Dynamik der vielfach divergierenden und konvergierenden Entdeckungspfade, welche engste Verknüpfungen zwischen Nervenfunktionen und molekularen Prozessen, zwischen Wahrnehmungsleistungen und der Aktivität spezifischer Neurone sowie zwischen kognitiven Leistungen und der Integrität spezifischer Hirnsysteme erkennbar machten, läßt aber auch erkennen, daß unser gegenwärtiger Kenntnisstand nur ein Durchgangsstadium ist, das bald Geschichte sein wird.
Lit.:Adelman, G.: Encyclopedia of Neuroscience. Amsterdam 1998. Arbib, M.A.: The Handbook of Brain Theory and Neural Networks. Cambridge 1995. Finger, S.: Origins of Neurosciences. Oxford 1994. Florey, E., Breidbach, O. (Hrsg.): Das Gehirn – Organ der Seele? Berlin 1993. Gazzaniga, M.S.: The Cognitive Neurosciences. Cambridge 1995. Gregory, R.L.: The Oxford Companion to the Mind. Oxford 1998. Kandel, E.R., Schwartz, J.H., Jessell, T.M.: Principles of Neural Science. New York 42000. Oeser, E.: Geschichte der Hirnforschung. Darmstadt 2002. Shepherd, G.M.: Foundations of the Neuron Doctrine. Oxford 1991. Shepherd, G.M.: Neurobiology. Oxford, New York 1997. Spillane, J.D.: The Doctrine of the Nerves. London, New York 1981. Squire, L.R.: The History of Neuroscience in Autobiography. San Diego 1998.
;)
Abb. 1: Diese Ganglienzellen aus der grauen Substanz des Rückenmarks eines Ochsen wurden von O. Deiters gezeichnet. Hier wird erstmals deutlich zwischen Dendriten und Axon unterschieden.
;)
Abb. 2: Der erste identifizierte Neuronentyp des Nervensystems: die von Purkinje 1837 gezeichneten großen Neurone des Kleinhirns, die seither seinen Namen tragen. Außerdem ist es die erste Darstellung der zellulären Struktur histologischer Schichten im Gehirn.
;)
Abb. 3: Anordnung von Neuronen und deren Verbindungsbahnen im Hippocampus auf der Grundlage Golgi-gefärbter Schnitte durch das Gehirn eines Nagetieres. Wie in vielen anderen seiner Zeichnungen deutete Ramón y Cajal hier die von ihm aus dem Prinzip der dynamischen Polarisierung der Neurone vermutete "nervöse Strömung", den Signalfluß, durch Pfeile an.
;)
Abb. 4: Die erste intrazelluläre Ableitung eines Aktionspotentials durch Hodgkin und Huxley (1939). Die hier an einem Riesenaxon eines Tintenfisches beobachteten Phänomene erwiesen sich von überraschender Generalität für fast alle Neurone in allen Tierstämmen. Links sind die Spannungsschwankungen in Millivolt ablesbar, unten sind die Zeitmarker durch jeweils 2 Millisekunden voneinander getrennt.
;)
Abb. 5: Rekonstruktion des Acetylcholinrezeptor-Kanalkomplexes eines elektrischen Fisches auf der Basis elektronenmikroskopischer Aufnahmen. Der Komplex hat einen Durchmesser von 8,5 nm und ragt deutlich in den Extrazellulärraum hinein. Der Pfeil zeigt auf die intrazelluläre Seite der Ionenpore (nach Toyoshima und Unwin, 1988).
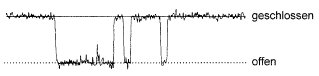
Abb. 6: Strom durch jeweils einen einzelnen Ionenkanal, der zwischen offenen und geschlossenen Zuständen hin und her springt. Der Unterschied der Leitfähigkeit beträgt ca. 2,5 pA, die Dauer der hier gezeigten Messung ca. 80 ms (nach B. Sakmann).
;)
Abb. 7: Darstellung einer wachsenden Nervenfaser in der Zellkultur im zeitlichen Abstand von 50 min; Harrison (1908) zeigte so erstmals Gestalt und Beweglichkeit des Wachstumskegels.
;)
Abb. 8: Zeichnung dendritischer Dornen mit ihren charakteristischen Köpfchen und Stielen an Purkinje-Zellen des Kleinhirns (Ramón y Cajal, 1904).
;)
Abb. 9: Das Muster der Expression von Homöobox-haltigen Genen (Hox und Krox) im Myelencephalon (My) der Maus. Mutationen in diesen zuerst in Drosophila entdeckten Gengruppen führen zur Übertragung der Strukturen eines Körpersegments auf ein anderes. Durch die segmentale Expression homologer Gene wurde ihre Rolle für die Spezifizierung von Körpersegmenten auch in anderen Tierstämmen einschließlich der Säuger bestätigt. Gp Grundplatte, Rh1–Rh8 Rhombomere, oV otische Vesikel, Rm Rückenmark, Mh Mittelhirn.
;)
Abb. 10: Die Regeneration des optischen Nerven nach Rotation eines Auges um 180 führt zu visuo-motorischen Reflexen, die um einen entsprechenden Winkel fehlgeleitet sind und bleiben. Mit diesen Versuchen zeigte Sperry 1956, daß auswachsende Nervenfasern ihr zentrales Zielgebiet unabhängig von der Geometrie der Wachstumsbedingungen oder der Sinneserfahrung finden können. Zusammen mit späteren Versuchen konnte Sperry den Beweis führen, daß Verhaltensleistungen unmittelbar auf spezifischen Mustern neuronaler Kontakte beruhen.
;)
Abb. 11: Der vollständige Zellstammbaum des Fadenwurms Caenorhabditis elegans: Unter den 959 Zellen des erwachsenen (hermaphroditischen) Organismus sind 358 Nervenzellen. Bei diesem Organismus haben Neurone mit gleichen Funktionen, z.B. Innervation der Bauchmuskulatur, das gleiche Teilungsschicksal. Während bei Wirbellosen das Zellteilungsschema stereotyp ist und kaum auf die spezifischen Umgebungsbedingungen reagiert, erfolgt die Entwicklung der Nervensysteme von Wirbeltieren unter massiver Einflußnahme von Umweltreizen.
;)
Abb. 12: Der erste Schritt zur Entdeckung des Nervenwachstumsfaktors gelang Levi-Montalcini 1951, als sie beobachtete, daß die Neurone eines explantierten sensorischen Ganglions eines Hühnchens zahlreiche Neuriten auswachsen lassen, wenn sich das Ganglion mit einem Stück Sarkom-Gewebe in einer gemeinsamen Kulturschale befindet.
;)
Abb. 13: Mit dieser Darstellung dokumentierte Sigmund Exner schon 1894 die Vorstellung, daß die Erzeugung präzisen Verhaltens nicht allein auf der Aktivität einzelner Neurone basiert, sondern ebenso von der spezifischen Architektur des zugrundeliegenden Nervennetzes abhängt.
;)
Abb. 14: Logische Verknüpfungen zwischen Nervenzellen. Mit den numerierten Dreiecken stellte McCulloch 1943 Nervenzellkörper dar, die Linien zeigen ihre Axone, die Verdickungen an ihren Enden erregende Synapsen, der Ring zeigt eine hemmende Synapse. Die neuronale Erregung fließt von den präsynaptischen Neuronen (links) zum postsynaptischen Neuron (rechts). Jede Synapse überträgt Aktivität im Wert von +1 (erregend) oder –1 (hemmend), die Schwelle des postsynaptischen Neurons soll 2 sein. Die 4 Zeichnungen stellen eine Erregungsübertragung (a), eine ODER-Schaltung (b), eine UND-Schaltung (c) und eine NICHT-Schaltung (d) dar.
;)
Abb. 15: Schema der hierarchischen Organisation von 3 Gehirntypen, die im Verlauf der Evolution der Säuger Bestandteil des Erbes des menschlichen Gehirns sind (nach P.M. MacLean, 1967).
;)
Abb. 16: Eine Ganglienzelle in der Netzhaut der Katze, deren Zellkörper und proximale Dendriten mit Hilfe des retrograden Transports eines axonalen Tracers angefärbt wurden, den die Zelle am Terminalgebiet ihres Axons im Colliculus superior aufgenommen hat. Die Entwicklung von Tracer-Techniken dieser Art war Voraussetzung dafür, die bestehenden Nervenbahnen im Gehirn zu kartieren und wesentliche Aspekte ihrer Organisation zu entschlüsseln.
;)
Abb. 17: Die Photographie des schwach vergrößerten Querschnitts durch das Gehirn einer Ratte zeigt die Verteilung (schwarze Bereiche) des Rezeptors für das Neuropeptid Substanz P. Die Darstellung der Rezeptoren erfolgt durch Aufbringen von Substanz P (Iod-125-markiert) auf die Hirnschnitte, die sich an die Rezeptoren bindet, und anschließenden autoradiographischen Nachweis des radioaktiven Markers (schwarze Stellen). (Nach R.B. Rothman, M. Herkenham, C.B. Pert).
;)
Abb. 18: Die Orientierungsspezifität von rezeptiven Feldern, wie sie zuerst von Hubel und Wiesel 1968 an Neuronen des primären visuellen Cortex der Katze beobachtet wurde. Ein Lichtbalken (linke Spalte), der über das rezeptive Feld (gestrichelter Kasten) des betreffenden Neurons hinweg bewegt wird, treibt das Neuron nur dann zu maximaler Aktivität (gemessen an der Anzahl der Aktionspotentiale, die rechts als senkrechte Striche erscheinen), wenn er die optimale Orientierung und Bewegungsrichtung (D) aufweist.
;)
Abb. 19: Die von Jasper und Penfield registrierten EEG-Kurven verschiedener Schlaf- und Wachzustände (Nach H.H. Jasper, N. Penfield, J.C. Erickson, 1941).
;)
Abb. 20: Diese Attrappenreize wurden von Tinbergen 1951 eingesetzt, um das Paarungs- und Attackierverhalten des Stichlings zu erforschen. Dabei zeigte sich, daß die exakte die Fischgestalt naturgetreu nachahmende Form der Attrappe (2. von oben) für die Auslösung artspezifischen Verhaltens weniger wirksam ist als eine rote Bauchseite, die nur der obersten Attrappe fehlt.








Wenn Sie inhaltliche Anmerkungen zu diesem Artikel haben, können Sie die Redaktion per E-Mail informieren. Wir lesen Ihre Zuschrift, bitten jedoch um Verständnis, dass wir nicht jede beantworten können.