Lexikon der Neurowissenschaft: Stickoxid-Synthase
Stickoxid-Synthasew, Stickstoffmonoxid-Synthase, NO-Synthase, Enitric oxide synthase (Abk. NOS), eine Familie von Enzymen, die (ATP-unabhängig [Adenosintriphosphat], daher keine "Synthetasen") die Bildung von Stickoxid (NO·) katalysieren ( siehe Abb. 1 ). Drei Klassen von NOS werden unterschieden, benannt nach dem Zell- bzw. Gewebetyp, aus dem sie zur näheren Charakterisierung erstmalig isoliert worden sind: neuronale NOS (nNOS, brain NOS; NOS I), Makrophagen-NOS (macNOS, NOS II), auch induzierbare oder immunologische NOS (iNOS genannt) und die endotheliale NOS (eNOS; NOS III). Die drei NOS-Isoformen haben eine ähnliche Domänenstruktur und sind mit der Cytochrom-P450-Reductase näher verwandt ( siehe Abb. 2 ). – Von allen NOS-Isoformen sind jeweils alternative Spleißformen bekannt (alternatives Spleißen). Neurophysiologisch besonders relevant ist die nNOS. Histochemisch gab es bereits seit 1964 Hinweise auf das Enzym in Form der Diaphorase-Reaktion, bei der die Oxidoreductase-Eigenschaft des Enzyms genutzt wird, um ein farbloses Tetrazoliumsalz (zumeist nitroblaues Tetrazoliumsalz, NBT) zu einem unlöslichen Farbstoff, dem Formazan, zu reduzieren. Die sogenannte NADPH-Diaphorase erwies sich später als weitgehend identisch mit NOS, indem Diaphorase-Histochemie und NOS-Immunohistochemie deckungsgleich einzelne Nervenzellen mit allen ihren Fortsätzen gegenüber der Masse nicht oder nur sehr schwach gefärbter Neuronen hochgradig markieren, vergleichbar einer Golgi-Imprägnation. nNOS und eNOS sind größtenteils konstitutiv exprimiert und durch den Calcium/Calmodulin-Komplex aktivierbar (Calcium, Calmodulin). Erhöhte intrazelluläre Calciumspiegel, wie sie z.B. aus einer Aktivierung von Glutamatrezeptoren vom NMDA-Typ resultieren, führen zum Anstieg der NO-Produktion. Das Vorkommen von eNOS im Gehirn außerhalb von Endothelzellen, z.B. in Neuronen, wurde zwar des öfteren behauptet, ist aber aufgrund neuerer Befunde eher unwahrscheinlich. Die iNOS wird normalerweise nicht im Gehirn exprimiert. Ihre Biosynthese läßt sich durch Entzündungsreize (experimentell bevorzugt durch bakterielles Lipoprotein und Gamma-Interferon) induzieren. Die iNOS-Aktivität ist wegen einer festen Kopplung des Enzymproteins mit Calmodulin calciumunabhängig. Typisch für die iNOS-Expression im Zentralnervensystem sind aktivierte Mikrogliazellen. Ihr Vorkommen in anderen Gliazellen-Typen oder in Neuronen ist noch strittig. – Das regionale und zelltypspezifische Auftreten der einzelnen NOS-Isoformen hat je nach der Rolle, die das NO in diesen Strukturen spielt, physiologisch wie pathophysiologisch eine sehr unterschiedliche Bedeutung. Studien an knock-out Mäusen haben im Falle der nNOS-Defizienz bislang (offenbar als Folge von Kompensationsvorgängen) nur geringfügige Auswirkungen gezeigt, u.a. erhöhte Aggressivität, verändertes Fortpflanzungsverhalten sowie, als Folge des Funktionsverlustes "nitrerger" Neurone im gastroenteralen Nervensystem, ein verengter Sphincter pylori und mithin ein auffällig geblähter Magen. Zugleich verringerte sich das Infarktvolumen bei experimenteller Ligatur von Hirngefäßen, was als Zeichen der Mitwirkung der nNOS bei neurodegenerativen Prozessen gedeutet wird. Über die PDZ-Domäne vermag sich die nNOS an die postsynaptische Dichtezone (PSD) zu binden und erhält so eine strukturelle Kopplung zu NMDA-Rezeptoren. Der durch die letzteren regulierte Calcium-Einstrom in die Zelle vermag vermutlich direkt die NO-Produktion zu erhöhen mit vermutlich unterschiedlichen Konsequenzen: Postsynaptisch generiertes NO kann über Diffusion zum präsynaptischen Element, also retrograd, die Transmissionsvorgänge modulieren (z.B. Langzeitpotenzierung, Langzeitdepression), NO kann aber auch im Falle einer excitotoxischen Erhöhung der Glutamatfreisetzung verstärkt gebildet und damit als suizidaler Faktor wirksam werden (Glutamatexcitotoxizität). – Alle NOS-Formen weisen Phosphorylierungsstellen auf, deren physiologische Bedeutung für die Regulation der NOS-Aktivität noch unklar ist. Experimentell ist NOS u.a. durch Derivate des Substrates Arginin hemmbar, z.B. Nitroarginin oder Nitroarginin-Methylester, aber auch durch 7-Nitroindazol, Aminoguanidin und Thioharnstoffderivate. Der Entwicklung NOS-Isoform-spezifischer Inhibitoren wird bei der Verfolgung von neuartigen Therapieansätzen große Bedeutung beigemessen.
G.W.
NO-Synthase-positive Neurone wurden in einer Vielzahl terrestrischer und mariner Mollusken nachgewiesen. Die immunhistochemische Lokalisation von NOS und die enzymhistochemische Detektion der NADPH-Diaphorase in Ganglien und peripheren Nerven von Tintenfischen weisen auf eine NO-Bildung und Freisetzung in diesen Strukturen hin. Im Zentralnervensystem dieser Tiere konnte NOS in den niederen motorischen Zentren, einschließlich der inferioren und superioren Loben (Freßzentren), in einigen höheren motorischen Zentren, im Lernzentrum (vertikaler, subvertikaler und superiorer frontaler Lobus) und im visuellen System nachgewiesen werden. Immunopositiv waren auch das olfaktorische System und das Epithel der Saugnäpfe. Stickoxid-Mechanismen spielen bei der Koordinierung von Lernvorgängen eine wesentliche Rolle. Durch den Einsatz des NOS-Inhibitors L-NAME (NG-Nitro-L-Arginin-Methyl-Ester) konnten spezifische Verhaltensmuster selektiv blockiert werden.
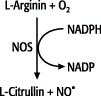
Stickoxid-Synthase
Abb. 1: von der Stickoxid-Synthase katalysierte Reaktion
;)
Stickoxid-Synthase
Abb. 2:Struktur der drei Stickoxid-Synthase-Formen und ihr Vergleich mit der Cytochrom-P450-Reductase
BH4 Tetrahydro-Biopterin; PDZ Anheftungsstelle für membranale PDZ-Domänen; Häm Bindungsstelle für Häm-Komplex; P Phosphorylierungsstelle; CaM Bindungsstelle für Calmodulin; FMN, FAD, NADPH, Bindungsstellen für Co-Enzyme; Myr Myristoylierungsstelle (zur Membranverankerung); TM Transmembrandomäne.







Wenn Sie inhaltliche Anmerkungen zu diesem Artikel haben, können Sie die Redaktion per E-Mail informieren. Wir lesen Ihre Zuschrift, bitten jedoch um Verständnis, dass wir nicht jede beantworten können.