Lexikon der Biologie: formative Teilung
formative Teilung, formative Zellteilung, Zellteilung (Cytokinese), bei der den entstehenden Tochterzellen ein unterschiedliches Entwicklungsschicksal zugewiesen wird ( vgl. Abb. 1 ). Der Begriff der formativen Teilung geht auf die von A. Weismann geprägte „erbungleiche Teilung“ zurück ( vgl. Infobox ). Weismanns Konzept wurde inzwischen dahingehend korrigiert, daß den Tochterzellen ein unterschiedlicher Satz sog. Entwicklungsdeterminanten zugeteilt wird, der dann eine unterschiedliche Differenzierung der Tochterzellen bewirkt. Die Natur dieser Entwicklungsdeterminanten ist noch weitgehend unverstanden – es könnte sich um unterschiedliche Transkriptionsfaktoren handeln, die in den Tochterzellen unterschiedliche Gene anschalten; es könnten unterschiedliche Proteine sein, die solche Transkriptionsfaktoren unterschiedlich aktivieren, und es wäre sogar denkbar, daß rein strukturelle Unterschiede der Tochterzellen eine Rolle spielen. Im Extremfall einer Mosaikentwicklung wird die Entwicklung durch eine streng festgelegte Folge von formativen Teilungen bestimmt (z.B. bei Caenorhabditis elegans). Formative Teilungen zeigen oft (aber nicht immer) Abweichungen vom normalen Ablauf einer Zellteilung. In der tierischen Entwicklung findet sich im Zusammenhang mit formativen Teilungen sehr oft eine abweichende Orientierung der Teilungsachse, in der pflanzlichen Entwicklung sind formative Teilungen zumeist asymmetrische Zellteilungen, bei denen Tochterzellen verschiedener Größe entstehen. Formative Zellteilungen lassen sich schon an Prokaryoten beobachten: bei der Teilung des gestielten Bakteriums Caulobacter crescentus (Caulobacter) entstehen 2 voneinander verschiedene Tochterzellen mit verschiedenen Entwicklungswegen ( vgl. Abb. 2a ): die gestielte, an der Basis gelegene Zelle wird zu einer Art Stammzelle, die immer wieder Tochterzellen abschnürt, während die spitzenwärts gelegene Tochterzelle Geißeln (Bakteriengeißel) bildet und frei beweglich ist. Nach einer freibeweglichen Phase wirft sie die Geißeln ab und bildet an deren Stelle einen Stiel aus. In diesem Fall ist die molekulare Grundlage der Entwicklungsdeterminanten bekannt. Es handelt sich um Proteine, die für die freibewegliche Lebensweise nötig sind und als Geißelanker bzw. als Rezeptoren für die Chemotaxis dienen ( vgl. Abb. 2c ). Diese Proteine werden in neugebildete Membranbereiche eingebaut – aufgrund des interkalaren Wachstums von Caulobacter also in einer gürtelartigen Zone rings um die Zellmitte. Kurz vor der Abschnürung der Tochterzelle werden diese Proteine über kurzlebige Proteasen abgebaut. Das Signal für die Proteasebildung scheint die abgeschlossene Verdopplung der DNA zu sein. Die Protease verschwindet daraufhin wieder, während die Bildung der Entwicklungsdeterminanten weiterläuft. Dadurch wird erreicht, daß nur die unmittelbar vor der Abschnürung gebildeten Membranbereiche, also die Zellpole, mit diesen Determinanten besetzt sind. Während der nachfolgenden Zellteilung durchläuft dieser Pol eine Reihe von Veränderungen (Abb. 2b). Zunächst dient er als Ankerstelle für die Bildung neuer Geißeln, und nach einer weiteren Zellteilung wächst von ihm der neue Stiel aus. Grundlage für die Steuerung der formativen Zellteilung ist in diesem Beispiel eine ausgeprägte Zell-Polarität. – Bei vielen „Blaualgen“ (Cyanobakterien) findet man Zellfäden, die aus blaugrünen, photosynthetisch aktiven Zellen und aus größeren, pigmentfreien Heterocysten aufgebaut sind. Diese Heterocysten sind zur Nitratassimilation (assimilatorische Nitratreduktion) in der Lage. Bei dem fädigen Cyanobakterium Anabaena entstehen die Heterocysten infolge einer asymmetrischen Teilung aus den photosynthetischen Zellen, wobei die Heterocyste sich aus der kleineren Tochterzelle entwickelt. Jedoch sind nicht alle Zellen eines Fadens zu einer solchen formativen Teilung in der Lage, sondern nur jene, die nicht zu nahe an schon bestehenden Heterocysten gelegen sind. Die schon gebildeten Heterocysten erzeugen in ihrer Umgebung also ein Hemmfeld unbekannter Natur, das weitere formative Teilungen verhindert. – Bei Tieren werden formative Teilungen vor allem während der frühen Embryogenese (Embryonalentwicklung) beobachtet. Während gewöhnlich die künftige Teilungsachse nach jeder Teilung um 90° weitergedreht wird, so daß die Tochterzellen kreuzartig angeordnet werden, werden formative Teilungen meist in derselben Achse angelegt wie die vorangegangene Zellteilung. Bei dem Fadenwurm Caenorhabditis elegans ist die erste Teilung der befruchteten Eizelle zwar symmetrisch, die beiden Tochterzellen verhalten sich jedoch sehr unterschiedlich während der nachfolgenden Zellteilung. Während in der einen Zelle (AB-Zelle) die Teilungsachse um 90° kippt, wird sie in der anderen Tochterzelle (P-Zelle) dagegen auf 180° weitergedreht. Kurz bevor sich die P-Zelle teilt, werden in der Zelle sog. P-Körperchen zum einen Zellpol transportiert, vermutlich entlang von Actin-Mikrofilamenten (Actinfilament). Die abweichende Teilungsachse hat nun zur Folge, daß nur eine der aus der P-Zelle entstehenden Tochterzellen diese P-Körperchen erbt und nun eine ganz andere Entwicklung einschlägt, wie die andere Tochterzelle. Bei diesen P-Körperchen handelt es sich also um Entwicklungsdeterminanten, vermutlich um untranslatierte mRNA. Es gibt nun sog. par-Mutanten, bei denen die abweichende Spindeldrehung in der P-Zelle unterbleibt, was zu einer falschen Plazierung der P-Körperchen und infolgedessen zu massiven Entwicklungsstörungen führt. – Bei Niederen Pflanzen hat sehr oft die erste Teilung einer Keimzelle formativen Charakter. Die erste, asymmetrische Teilung von Zygoten des Blasentangs Fucus (Fucales) trennt eine kleinere, von dichtem Plasma erfüllte Spitzenzelle von einer größeren, stärker vakuolisierten Basalzelle. Die Spitzenzelle entwickelt sich zum photosynthetisch aktiven Thallus, die basale Zelle zum Rhizoid. Die Achse dieser ersten Zellteilung kann durch Umwelteinflüsse wie Blaulicht, Schwerkraft (Gravitationsbiologie) und elektrische Felder ausgerichtet werden. Durch eine Bestrahlung mit starkem, polarisiertem Blaulicht läßt sich eine symmetrische Teilung hervorrufen. Aus den beiden nun gleich großen Tochterzellen entsteht dann jeweils ein Rhizoid. Sporen des FarnsCeratopteris (Parkeriaceae) teilen sich in ähnlicher Weise, wobei hier die Teilungsachse durch die Schwerkraft festgelegt wird. Nach Behandlung mit Mikrotubuligiften wird diese Teilung symmetrisch. In diesem Fall entsteht aus den beiden nun gleich großen Tochterzellen jeweils ein Thallus. Diese Beobachtungen legen nahe, daß es sich hier um formative Teilungen handelt. – Bei Höheren Pflanzen ist die erste Teilung der Mikrospore im Pollenkorn (Pollen) asymmetrisch. Wird sie durch Behandlung mit Kälte oder Mikrotubuligiften (Mitosegifte) symmetrisch gemacht, beobachtet man oft die Bildung eines abnormen (haploiden) Embryos anstelle der normalen Bildung eines Pollenschlauchs. Ebenso ist die erste Teilung der Zygote im mütterlichen Embryosack asymmetrisch und trennt die Vorläuferzelle von Embryo und Suspensor (Embryoträger). Bei der Mutante Gnom (von Arabidopsis; Schmalwand) ist diese erste Teilung jedoch symmetrisch, und dies ist mit einem Ausfall der Entwicklung der Wurzel (Wurzelbildung) verbunden. Formative Teilungen wurden auch bei der Bildung der verschiedenen Wurzelgewebe vermutet – im Meristem (Bildungsgewebe) kippt bei sog. Cortex-Vorläuferzellen die Teilungsachse von antiklin nach periklin, wodurch eine äußere und eine innere Tochterzelle entsteht. Die Nachkommen der inneren Tochterzelle differenzieren sich zur Endodermis, während die Nachkommen der äußeren Tochterzelle zu Cortexzellen (Rinde) werden. Mit Hilfe eines Laserstrahls (Laser) können nun einzelne dieser Endodermiszellen entfernt werden, worauf die benachbarten Cortexzellen in die entstehende Leerstelle einwandern. Die eingewanderten Cortexzellen differenzieren sich dann zu Endodermiszellen. Sie entwickeln sich nun also nicht gemäß ihrer Herkunft, sondern gemäß ihrem neuen Ort. Das Entwicklungsschicksal pflanzlicher Zellen wird also weniger durch ihre Herkunft bestimmt, sondern durch Signale, die sie von ihren Nachbarn empfangen. Bei Höheren Pflanzen scheint also die Bedeutung der formativen Teilung stark eingeschränkt zu sein. asymmetrische Teilung.
P.N.
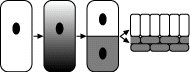
formative Teilung
Abb. 1: Konzept der formativen Teilung. In der Mutterzelle existiert ein Gefälle von Entwicklungsdeterminanten, die bei der Teilung ungleich auf die Tochterzellen verteilt werden. Die Nachkommen der beiden Tochterzellen schlagen dann unterschiedliche Entwicklungswege ein.
;)
formative Teilung
Abb. 2: Formative Zellteilung bei dem Bakterium Caulobacter crescentus:a Entwicklungskreislauf; b Verhalten der beiden Zellpole während der Teilung; der neu angelegte Pol durchläuft im Lauf der Entwicklung verschiedene Reifezustände. c Neuanlage des Geißelpols







Wenn Sie inhaltliche Anmerkungen zu diesem Artikel haben, können Sie die Redaktion per E-Mail informieren. Wir lesen Ihre Zuschrift, bitten jedoch um Verständnis, dass wir nicht jede beantworten können.