Lexikon der Geowissenschaften: Fische
Fische, aquatische, kaltblütige Wirbeltiere, die mit Hilfe von Kiemen atmen. Systematisch teilt man die Fische in mehrere Klassen, wobei grundsätzlich die kieferlosen Fische (Agnatha) von den kiefertragenden Fischen (Gnathostomata) unterschieden werden ( Abb. 1 ). Diese beiden Hauptgruppen haben sich bereits im Kambrium getrennt. Im Fossilbericht sind bisher keine Zwischen- oder Übergangsformen bekannt geworden. Auch der grundsätzlich verschiedene Bau des Kiemenapparates macht verwandtschaftliche Beziehungen von kieferlosen und kiefertragenden Fischen sehr unwahrscheinlich.
Der Ursprung der Fische liegt innerhalb der Chordatiere (Chordata). Neben den Wirbeltieren (Vertebrata) werden zu dieser auch die Hemichordata (Eichelwürmer), die Urochordata (Manteltiere = Tunicata) und die Cephalochordata (z.B. Branchiostoma, das Lanzettfischchen) gezählt. In allen diesen Gruppen treten (z.T. nur in den Larval- oder Embryonalstadien) Organisationsmerkmale wie ein versteifendes, rohrförmiges Stützorgan (Chorda dorsalis), ein dorsal gelegenes Neuralrohr sowie Kiemenspalten und V-förmige Muskelsegmente (Myomere) auf, die anderen Tierstämmen fehlen. Ein kleines, nur etwa 5 cm langes Fossil aus dem berühmten mittelkambrischen Burgess Shale in British Columbia, Kanada, gilt als der älteste Chordate: Die fischchenförmige Pikaia ( Abb. 2 ) zeigt eine als Chorda interpretierte Struktur sowie segmentierte Muskeln. Neue Funde von Agnathenresten aus dem chinesischen Unterkambrium machen dem bisher als ältestes Wirbeltier angesehenen Anatolepis Konkurrenz, das aus dem Oberkambrium Wyomings (USA) stammt. Die überlieferten kleinen Schuppenteile mit einer charakteristisch genoppten Oberfläche bestehen aus biologischem Apatit, dem Baustoff von Knochen und Zähnen der Vertebrata. Zwischen diesen frühesten Wirbeltierresten und der diversen Fischfauna aus dem Zeitbereich Silur/Devon ("Zeitalter der Fische") ist der Fossilbericht sehr spärlich.
Die Agnatha, in älterer Literatur auch als Ostracodermen bezeichnet, erscheinen im oberen Kambrium und entwickeln im Silur und Devon eine erstaunliche Formenfülle. Die allermeisten Gruppen kieferloser Fische sterben jedoch zum Ende des Devons wieder aus, wobei sicherlich der Konkurrenzdruck der sich nun stark diversifizierenden kiefertragenden Fische eine entscheidende Rolle gespielt hat. Rezent sind nur noch die Petromyzontia und die Myxinoidea vorhanden. Im Devon des Baltischen Schildes dienen die Agnatha als Zonenfossilen. Morphologisch zeichnen sich die fossilen Vertreter der kieferlosen Fische durch einen kräftigen Knochenpanzer oder zumindest ein knöchernes Schuppenkleid aus, das große Teile des Körpers bedeckte.
Eine der drei großen Gruppen innerhalb der Agnatha sind die Osteostraci. Im Fossilbericht erscheinen sie im Obersilur und haben meist einen dorsoventral abgeplatteten Körper und einen asymmetrischen Ruderschwanz zur Fortbewegung. Kennzeichnende Merkmale dieser Gruppe sind paarige Flossen und ein knöchernes Innenskelett im Schädelbereich, wo elektrische Sinnesorgane vermutet werden. Ökologisch werden die Osteostraci als träge, bodenbewohnende Strudler gedeutet, die sowohl in küstennahen marinen Sedimenten als auch in Süßwasserablagerungen vorkommen. Eine im Silur und Devon, vorwiegend im Süßwasser auftretende Gruppe sind die spindelförmigen Anaspida, die als aktive Schwimmer betrachtet werden. Ihr Besatz mit Knochenschuppen wird von stratigraphisch älteren zu jüngeren Formen zunehmend abgebaut. Aus ihnen könnten die rezenten Neunaugen hervorgegangen sein. Die Heterostraci sind die älteste Gruppe innerhalb der kieferlosen Fische. Sie erscheinen bereits im Kambrium und weisen in der vorderen Körperhälfte eine kräftige Knochenpanzerung auf, während der hintere Körper mit Knochenschuppen bedeckt ist. Von der vierten Agnathen-Gruppe, den Coelolepida, ist nur wenig bekannt, da sie ihre Knochenpanzerung bis auf kleine, in der Haut gelegene Schüppchen reduziert haben und daher fossil nur schlecht erhaltungsfähig sind. Die Galeaspida mit ihren bizarren Körperformen sind vom Untersilur bis Mitteldevon in China und Nordvietnam verbreiteten und haben sich dort vermutlich isoliert entwickelt. Eine weitere Gruppe eigenartig gestalteter Fische sind die erst in den letzten Jahren beschriebenen Thelodontier, die vom oberen Ordovizium bis zum Ende des Devons nachgewiesen sind.
Im unteren Silur treten im Fossilbericht die ersten kiefertragenden Fische (Gnathostomata) auf, die in die ausgestorbenen Placodermen (Panzerfische), die Chondrichthyes (Knorpelfische), die Osteichthyes (Knochenfische) und die eigentümlichen Acanthodii ("Stachelhaie") gegliedert werden ( Abb. 1 ). Der Erwerb von Kiefern stellt innerhalb der Wirbeltiere einen einschneidenden Evolutionsschritt dar. Er ebnete den Weg zu einer räuberischen Ernährungs- und Lebensweise und damit die Radiation in bis dahin von Fischen unbesetzte ökologische Nischen. Die Kiefer entwickelten sich aus transformierten Kiemenbögen: Mit der Ausdehnung der Mundhöhle gingen die vordersten zwei bis drei Kiemenbögen verloren, während sich das nachfolgende Paar vergrößerte und zu Kiefern umbildete.
Die Placodermen sind oft sehr eigenartig gestaltete Fische mit einem massiven, panzerartigen Exoskelett aus Knochenplatten. Sie gelten als die primitivsten kiefertragenden Wirbeltiere. Die meisten Panzerfische sind marin, die Vertreter der Ordnung Antiarchi ( Abb. 3a ) kommen jedoch überwiegend in Süßwasserablagerungen vor. Placodermen treten erstmals an der Wende Silur/Devon auf und haben ihre Blütezeit während des Devons. Bereits im Unterkarbon sterben sie ohne rezente Nachkommen aus. Placodermen besaßen einen starren, unbeweglichen Unterkiefer, so daß zum Öffnen des Mauls der gesamte Kopf gehoben werden mußte. Daher waren Kopf- und Rumpfpanzer der Placodermen über ein Nackengelenk beweglich miteinander verbunden. Vermutlich waren nur diejenigen Formen gute Schwimmer, die eine vergleichsweise geringe Panzerung aufwiesen (z.B. Coccosteus, Abb. 3b ). Vor allem innerhalb der Ordnung Arthrodira gab es aktive Räuber mit furchterregenden Dimensionen, wie z.B. Dunkleosteus, der über 10 m lang werden konnte.
Das Skelett der Chondrichthyes ist aus Knorpelgewebe aufgebaut, weshalb sie ein wesentlich schlechteres Fossilisationspotential haben als Fische, deren Stützorgane aus Knochen bestehen. Die Chondrichthyes gliedern sich zum einen in die Elasmobranchii, zu denen die rezenten Haie (Selachii, Abb. 4 ) und Rochen (Batomorpha, Abb. 5 ) sowie deren fossile Vertreter gehören. Zum anderen zählen die bizarren Chimären (Holocephali) zu dieser Klasse. Entgegen älterer Annahmen stammen die Chondrichthyes nicht von den Placodermen ab. Vor allem durch Gemeinsamkeiten im Kieferbau und beim Zahnersatz lassen sie sich mit den Osteichthyes (Knochenfischen) von einem gemeinsamen, fossil bisher nicht belegten Vorfahren herleiten. Bereits im Untersilur zeigen die Haie eine rasche Radiation in viele Familien. Die meisten Entwicklungslinien erlöschen jedoch an der Perm/Trias-Grenze, lediglich die Hybodontoidea bestehen weiter bis ins Alttertiär fort. Eine zweite Radiationsphase erleben die Elasmobranchii im älteren Mesozoikum. Vermutlich aus den Ctenacanthoidea gehen die modernen Haie (Neoselachii) hervor, und im unteren Jura Norddeutschlands können die frühesten bekannten Rochen nachgewiesen werden. Bereits in der Kreide sind die Elasmobranchier weitgehend in die rezent vorkommenden Gruppen differenziert. Gemeinsames Merkmal aller Chondrichthyes ist das knorpelige Endoskelett, das bei erwachsenen Exemplaren auch prismatisch calcifizierten Knorpel enthalten kann. Die männlichen Tiere haben ein äußeres, paariges Kopulationsorgan, die Pterygopodien, die sich aus einem Teil der Bauchflossen differenziert haben ( Abb. 4a ). Es dient der bei Fischen ungewöhnlichen inneren Befruchtung. Fast alle Formen sind marin, nur wenige Arten sind Süßwasserbewohner. Die Außenhaut der Elasmobranchii ist mit winzigen schmelzüberzogenen Hautzähnchen bedeckt, die im Bautyp den "echten" Zähnen homolog sind. Diese auch fossil nachgewiesenen Placoidschuppen setzen den Wasserwiderstand beim Schwimmen beträchtlich herab. Einige Haie haben Stacheln an Flossen oder Kopf ausgebildet. Die charakteristischen, meist mehrspitzigen und/oder mit Sägekanten versehenen Haizähne ( Abb. 4b-e ) werden im Rhythmus von meist wenigen Wochen ersetzt. Bedingt durch diesen raschen Zahnwechsel sind die mancherorts massenhaft überlieferen Haizähne die häufigsten Chondrichthyer-Reste im Fossilbericht. Viele der modernen und fossilen Haie sind torpedoförmig gebaut und haben eine räuberische Lebensweise. Daneben gibt es aber immer Formen, die durophag sind, d.h. sich mit flachen Knackzähnen ( Abb. 4f ) von Mollusken ernähren. Rochen sind dorsoventral stark abgeflacht, zeigen besonders vergrößerte, flügelartige Brustflossen, wohingegen die Flossen der Bauch- und Schwanzregion stark reduziert sind ( Abb. 5 ). Ihre Lebensweise ist meist bodenbezogen, und mit ihren Quetschzähnen ernähren sie sich bevorzugt von hartschaligen Meeresorganismen.
Eine Gruppe niederer Gnathostomata mit unsicherer systematischer Zuordnung sind die Acanthodier. Sie weisen sowohl auf der Dorsal- als auch auf der Ventralseite des Körpers kräftige Stacheln auf ("Stachelhaie"). Diese meist räuberisch lebenden Fische sind bereits im Untersilur nachgewiesen und haben ihre Blütezeit im Devon. Aus permischen Süßwasserablagerungen gibt es jedoch auch zahnlose Gattungen (z.B. Acanthodes), die ökologisch als Strudler interpretiert werden. Die Acanthodier sterben ohne rezente Nachfahren im Perm aus.
Die Osteichthyes (Knochenfische) sind seit dem Obersilur durch Schuppen belegt und treten damit sehr früh im Fossilbericht der Vertebraten auf. Sie sind charakterisiert durch ein verknöchertes Innenskelett und die Ausbildung einer Schwimmblase als hydrostatisches Organ. Dies macht sie fossil und rezent zur erfolg- und formenreichsten Fischklasse. Die Actinopterygii teilen sich zum einen in die altertümlichen Chondrostei, die im Paläozoikum ihre Blütezeit hatten, und zum anderen in die Neopterygii. Der überwiegende Teil der heutigen Fische gehört in letztere, gegen Ende des Paläozoikums erscheinende Gruppe. Aus ihr entwickeln sich in der Obertrias die ersten Teleostier. Im Jura und in der Kreide sind v.a. die primitiveren Teleostier verbreitet. Die Euteleostier, zu denen von der Forelle bis zum Seepferdchen alle modernen Knochenfische gehören, dominieren ab dem Tertiär. Für den Stammbaum der Wirbeltiere sind jedoch die Sarcopterygii das wichtigere Taxon innerhalb der Knochenfische, da sich aus ihnen die landlebenden Tetrapoden entwickelt haben. Zu den Fleischflossern zählen die Dipnoi (Lungenfische), die Coelacanthiformes sowie die paläozoischen Rhipidistia. Gemeinsame Merkmale sind v.a. die namengebenden fleischigen Flossen sowie innere Nasenöffnungen (Choanen). Die frühesten Lungenfische sind aus dem Unterdevon Chinas bekannt, in der rezenten Fauna sind sie nur noch mit drei Gattungen vertreten. Die Dipnoi ( Abb. 6a ) sind gekennzeichnet durch Spezialisierungen im Schädelbau und die lungenartige Ausstülpung ihres Darmes. Die Coelacanthiformes erscheinen im Mitteldevon in Süßwasserablagerungen, sind aber ab der Trias auch im marinen Bereich heimisch. Als wissenschaftliche Sensation wurden Ende der 30er Jahre rezente Exemplare dieser seit der Oberkreide als ausgestorben geglaubten Gruppe als Latimeria beschrieben ( Abb. 6b ). Aus den Rhipidistiern sind im Oberdevon die Amphibien hervorgegangen. Die Baupläne beider Gruppen zeigen viele Gemeinsamkeiten: Insbesondere die paarigen Rhipidistierflossen sind bereits in diejenigen Knochenelemente differenziert, die eine Tetrapodenextremität ausmachen. Die anatomisch gut bekannte, oberdevonische Gattung Eusthenopteron ( Abb. 7 ) sowie die weniger vollständig überlieferte Gattung Panderichthyes gelten zur Zeit als die besten Modelle derjenigen Fischgruppe, die an der Schwelle zwischen Wasser und Land steht. [DK]
Literatur: [1] BENTON, M.J. (1997): Vertebrate Palaeontology. – London u.a. [2] CARROLL, R.L. (1993): Paläontologie und Evolution der Wirbeltiere. – Stuttgart/New York. [3] LONG, J.A. (1995): The rise of fishes: 500 million years of evolution. – Baltimore/London. [4] ROMER, A.S. & PARSONS, T.S. (1991): Vergleichende Anatomie der Wirbeltiere. – Hamburg/Berlin.
Fische 1: stratigraphisches Auftreten und relative Häufigkeit der verschiedenen Fischformen. Die Agnathen sind vorwiegend paläozoisch verbreitet, und rezent nur noch mit Petromyzon und Myxine vertreten. Alle großen Gruppen der kiefertragenden Fische sind ab dem Devon nachweisbar, ausgestorben sind die Placodermen und die Acanthodii. Die dominierenden Fische sowohl im marinen Bereich als auch im Süßwasser sind heute die Teleostier, die zu den Strahlenflossern gehören. Aus den Rhipidistiern sind im Oberdevon die landlebenden Tetrapoden hervorgegangen. Fische 1:
;)
Fische 2: Rekonstruktion des ältesten bekannten Chordaten, Pikaia gracilens, aus dem mittelkambrischen Burgess Shale (Kanada). Das Vorderende weist nach rechts, Länge ca. 5 cm.
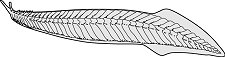
Fische 3: Vertreter der Placodermen (Panzerfische): a) Bothriolepis (Ordnung Antiarchi). Der massive Kopfpanzer und Rumpfpanzer sind über ein Nackengelenk beweglich miteinander verbunden, Länge ca. 40 cm; b) Coccosteus cuspidatus (Ordnung Arthrodira), eine schwächer gepanzerte Form, Länge ca. 35 cm. Fische 3:
;)
Fische 4: Vertreter der Chondrichthyes (Knorpelfische): a) mesozoische Haigattung Hybodus. Beachtenswert sind die kräftigen Flossenstacheln sowie die paarigen Kopulationsorgane (Pteryopodien); b-f) Beispiele unterschiedlicher Morphologien bei Haizähnen: b) Palaeohypotodus, Untereozän, c) Palaeocarcharodon, Paläozän, d) Eonotidanus, Unterkreide, e) Chlamydoselachus, rezent, Nordatlantik, f) Quetschzahn von Hylaeobatis, Unterkreide. Größen nicht maßstäblich. Fische 4:
;)
Fische 5: Vertreter der Batomorpha (Rochen): a) Cyclobatis, Oberkreide; b) schematische Darstellung des Kiefers und der Bezahnung von Mylobatis. Gut erkennbar ist die regelmäßige Anordnung der einzelnen Quetschzähne zu Reihen. Fische 5:
;)
Fische 6: Vertreter der Sarcopterygii (Fleischflosser, Osteichthyes): a) Epiceratodus (rezenter australischer Lungenfisch, Dipnoi); b) Latimeria (rezenter Quastenflosser, Coelacanthidae, Indischer Ozean). Größen nicht maßstäblich. Fische 6:
;)
Fische 7: Skelettrekonstruktion der oberdevonischen Rhipidistiergattung Eusthenopteron. Der Becken- und Schultergürtel zeigen bereits die Differenzierung in Ober- und Unterschenkel bzw. Ober- und Unterarm, wie sie für Tetrapoden üblich ist. Fische 7:
;)








Wenn Sie inhaltliche Anmerkungen zu diesem Artikel haben, können Sie die Redaktion per E-Mail informieren. Wir lesen Ihre Zuschrift, bitten jedoch um Verständnis, dass wir nicht jede beantworten können.